Academia.edu no longer supports Internet Explorer.
To browse Academia.edu and the wider internet faster and more securely, please take a few seconds to upgrade your browser.
2004, Journal of Zoology
https://doi.org/10.1017/S0952836904005011…
18 pages
Two hundred and fourteen skulls of Asian deer species (Cervini) were measured and the resulting craniometric data analysed using multivariate statistics. Our results suggest that elements of skull shape as determined by morphometrics can be used as phylogenetic characters and depict phylogenetic relationships among Cervini deer species that is not far removed from the picture given by molecular studies and other characters. Morphometricsbased phylogeny was used in combination with other phylogenies and palaeoenvironmental reconstructions to investigate factors that may have influenced evolutionary changes. Our data indicate the need for a revision of the genus and subgenus level taxonomy of the Cervini.

AI
The study finds that Elaphurus is craniometrically distinct from other Cervini due to significant length and width measurements, marking a high correlation with the first principal component of skull morphology.
Research demonstrates that cranial measurements can act as reliable phylogenetic indicators, as evidenced by a 82.4% accuracy in discriminant analysis classifying species groupings.
Antlered deer originated in the Middle to Late Tertiary, with key divergences during the Pleistocene, notably as early as 12.5-9.5 Ma for the three-pronged deer groups.
Environmental shifts from humid forests to open grasslands during the Pliocene and Pleistocene, combined with ancient hybridization events, likely fueled diversification within Cervini.
Evidence indicates Elaphurus may have a hybrid origin involving a Cervus ancestor, with controlled breeding experiments yielding over 300 viable hybrids with Cervus elaphus in captivity.








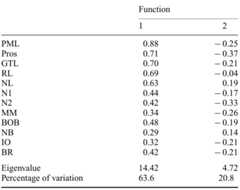
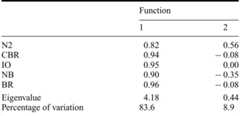









Earth, 2022
The article describes the paleobiogeographic history of the modern subfamilies so-called "crown deer" of the family Cervidae (Artiodactyla, Mammalia) in the world from the late Miocene to the late Pleistocene. The study overviews the taxonomic diversity and evolutionary radiation of Cervidae from all zoogeographic realms where this systematic group is present in the paleontological record. The evolutionary diversification of the fossil Cervidae is based on the estimations of species body masses that are regarded here as a proxy of occupied ecological niches. The study reveals two important evolutionary radiations of Cervidae during the late Miocene of Eurasia that gave the origin of the modern subfamilies Cervinae and Capreolinae. The evolutionary radiation of Capreolinae during the Pleistocene in South America shows a range of diversity comparable to the late Miocene radiations of Old World deer and provides multiple examples of evolutionary convergences with Eurasian Pleistocene cervids. The article discusses factors that shaped the modern biogeographic distribution of representatives of the subfamilies Cervinae and Capreolinae.
Scientific reports, 2017
Deer are an iconic group of large mammals that originated in the Early Miocene of Eurasia (ca. 19 Ma). While there is some consensus on key relationships among their members, on the basis of molecular- or morphology-based analyses, or combined approaches, many questions remain, and the bony labyrinth has shown considerable potential for the phylogenetics of this and other groups. Here we examine its shape in 29 species of living and fossil deer using 3D geometric morphometrics and cladistics. We clarify several issues of the origin and evolution of cervids. Our results give new age estimates at different nodes of the tree and provide for the first time a clear distinction of stem and crown Cervidae. We unambiguously attribute the fossil Euprox furcatus (13.8 Ma) to crown Cervidae, pushing back the origin of crown deer to (at least) 4 Ma. Furthermore, we show that Capreolinae are more variable in bony labyrinth shape than Cervinae and confirm for the first time the monophyly of the O...
Quaternary
The article presents a preliminary morphological description of the holotype of Megaloceros giganteus (Blumenbach, 1799) that serves for the description of the species. The article proposes a taxonomical and morphological revision of the nominotypical subspecies M. giganteus giganteus and morphological comparison with other subspecies of M. giganteus. The cluster analysis of diagnostic craniodental and antler characters revealed the systematic position and phylogenetic relationships of M. giganteus with other cervid groups. The genus Praedama is regarded as a closely related phylogenetic branch that linked to the direct cursorial forerunner of Megaloceros that evolved in the middle latitudes of Western Siberia and northern Kazakhstan. The genus Dama has a distant relationship with Megaloceros and represents an earlier phylogenetic branch that evolved in the Ponto-Mediterranean area. The article discusses the secondary adaptations of M. giganteus forms to forest and woodland habitats...
Journal of morphology, 2016
Deer (Cervidae) have a long evolutionary history dating back to the Early Miocene, around 19 million years ago. The best known fossils to document this history belong to European taxa, which all bear cranial appendages more or less similar to today's deer antlers. Despite the good fossil record, relationships of the earliest stem deer and earliest crown deer are much debated. This hampers precise calibration against the independent evidence of the fossil record in molecular clock analyses. While much has been written on the Early and Middle Miocene deer, only two phylogenetic analyses have been performed on these taxa to date mostly based on cranial appendage characters. Because the petrosal bone and bony labyrinth have been shown to be relevant for phylogeny in ruminants, we describe for the first time these elements for four iconic early cervids from Europe (Procervulus dichotomus, Heteroprox larteti, Dicrocerus elegans and Euprox furcatus) and include them in a phylogenetic a...
Quaternary International, 2008
Giant deer with palmate brow tines were placed either in a single genus, or those from west Eurasia were placed in Megaloceros and those from the east in Sinomegaceros, which implies local evolution. In order to see whether dispersals between the two areas may have occurred, we studied the morphology and interpreted the phylogeny of these deer.
Four dentary fragments of the deer species, Cervus cf. rewati (Cervidae, Artiodactyla, Mammalia) are described from the Dhok Pathan Formation, Middle Siwaliks (Siwalik Group, Pliocene) of northern Pakistan. The specimens under study include the lower teeth and exhibit the basic characters of Cervus rewati. Arif et al. (1991) described this species for the first time from the Upper Siwaliks (Pleistocene) but our findings extend the range of this species from the Pliocene of the Middle Siwaliks to the Upper Si-waliks of the Indian subcontinent. Es werden vier Unterkieferfragmente von der Cervus cf. rewati (Cervidae, Artiodactyla, Mammalia) aus der Dhok Pathan Forma-tion, Mittlere Siwaliks (Siwalik-Gruppe, Pliozän) aus dem nördlichen Pakistan beschrieben. Die Unterkieferzähne zeigen die prinzi-piellen Merkmale von Cervus rewati. Arif et al. (1991) beschrieben diesen Art zum ersten Mal aus den oberen Siwaliks (Pleistozän) und unsere Ergebnisse können diesen Art nun zum ersten Mal auch a...
Russian Journal of Genetics, 2005
Sequences of mitochondrial genes 12S and 16S rRNA (2445 bp) and the region of the nuclear betaspectrin gene (828 bp) were analyzed in members of the family Cervidae and in some other artiodactyls. Several molecular synapomorphies characteristic both of Cervidae and musk deer have been found. According to our data, Cervidae is a sister clade to Bovidae, which are very close to Moschidae. The family Giraffidae is exterior to this common clade, while Antilocapridae occupies a more basal position. The family Cervidae proper splits into three clades including the genera Cervus and Muntiacus (1), Capreolus, Hydropotes, Alces (2), and Rangifer, Odocoileus, and the remaining genera (3). In general, our phylogenetic reconstructions conform to the results of earlier molecular genetic studies, but substantially differ from the traditional taxonomy of Ruminantia.
Transactions of the Nebraska Academy of Sciences, 1985
Antlers of New World deer (Cervidae), though of considerable taxonomic importance, are rarely found in deposits earlier than Late Pleistocene. The one reasonably well known Blancan deer, Bretzia pseudalces, whose antlers are known from deposits in Washington and California, is an odocoilene, closely related to Odocoileus. An analysis of antler form and distribution patterns suggests that the immigration and divergence of the New World Odocoileinae (excluding Alces and Rangi!er) occurred in the latest Hemphillian and Early Blancan. Evolution of complexity in antler form in this group begins with a relatively simple pattern which lacks a brow tine immediately adjacent to the burr, possesses a single anteriorly directed tine, and has a larger (sometimes forked) posterior tine. This pattern is seen in Ozotoceras and in Capreolus. Bretzia altered this pattern by spreading the antlers laterally and by developing palmation of the posterior branch. The evolution of antlers of Odocoileus seems to begin with development of a helical form incorporating beam and anterior tine. The latter form appeared by the end of the Early Blancan, when Odocoileus apparently replaced Bretzia in the western parts of North America.
The primitive small-sized representatives of the "Cervus elaphus group" (Cervus nestii and Cervus abesalomi) appeared in Western Eurasia (Georgia and Italy) in Early Pleistocene. C. nestii already shows the basic primitive plane of antler morphology of the elaphus type and the peculiar elaphine cranial morphology. C. abesalomi Kahlke 2001 from Dmanisi (Georgia) seems to be closely related to C. nestii, but some-what more primitive in morphology of dentition. The Villafranchian Cervus from Italy and Georgia may not represent the direct ancestral form of the “elaphus” group. Perhaps, the first expansion of Cervus s.s. in Europe took place during the middle-late Villafranchian. The first elaphine immigrant C. nestii could give a start for radiation of various forms in the Mediterranean area that characterised by antlers with a single brow tine and unusually branched or palmed distal portions. Those endemic descent forms are C. elaphus aretinus Azzaroli with branched distal part of antler and very long braincase (Azzaroli, 1961; Di Stefano & Petronio, 1992) and C. elaphus palmidactyloceros with palmed antlers (Abbazzi, 1995). The Modern fauna of Western Mediterranean still contains a peculiar form of red deer both from morphological and from genetic points of view. This is a rather small-sized C. elaphus barbarus with primitive antlers similar to C. nestii and white spots on its body. Ludt et al. (2003) obtained a surprisingly high genetic differentiation of the Atlas stag together with the Corsican deer from the rest subgroups of the modern red deer with the time of divergence cca. 2.2 Ma (since the Middle Villafranchian). Such a significant time of divergence was taken with caution as a possible overestimation (Ludt et al., 2003). However, the archaic morphology of Atlas and Corsican deer and their genetic segregation may be regarded as an evidence for a long-time isolation of primitive red deer in the Atlas refugia. Therefore, one can assume that the Atlas stag is a Villafranchian survivor derived from C. nestii.
Iranian Journal of Veterinary Medicine, 2021
BACKGROUND: Persian fallow deer (PFD), Dama dama mesopotamica, is one of the rarest members of the Cervidae family currently listed as endangered by the International Union for Conservation of Nature. OBJECTIVES: Morphological variations in the skulls of male and female PFDs were evaluated in this investigation. METHODS: In cooperation with the Department of Environment, skulls and mandibles were obtained from five male and four female animals. After the usual practices of bone cleaning, the specimens were evaluated for morphological differences. Next, 29 parameters were measured on the skulls and mandibles by a digital Vernier caliper for morphometric studies. In addition, ten, six, and nine landmark points were defined on the left lateral photos of mandibles, dorsal, and left lateral photos of skulls, respectively. The points were digitized on twodimensional images using the TpsDig2 software. The shape differences between the two genders were analyzed using discriminate function analysis in the MorphoJ software. RESULTS: The interfrontal ridge was more prominent in male PFDs than in female animals. Moreover, there were some significant differences in the measured parameters, mostly in the mandible. The geometric morphometric evaluations showed no significant differences between the two genders. CONCLUSIONS: The findings of the present investigation revealed some morphological differences between the skulls of male and female PFDs.

Loading Preview
Sorry, preview is currently unavailable. You can download the paper by clicking the button above.
Fossil Studies, 2024
This study employs Principal Component Analysis to examine cranial measurements from both extant and fossil specimens spanning 38 species and comprising over 300 individuals within the subfamily Cervinae. Our findings elucidate craniometric distinctions among cervids characterized by varying body sizes and certain evolutionary adaptations. While our results generally corroborate previous assertions regarding the limited craniometric variability among plesiometacarpal deer, they also unveil specific cranial adaptations within certain genera and species. The Principal Component Analysis of craniometric data revealed that giant and large-sized deer display significantly broader ecomorphological diversity in cranial shape compared to small-sized deer. Secondly, small-sized deer exhibit greater uniformity in their cranial shape, appearing densely clustered on the factorial map. Thus, we conclude that body size imposes ecological constraints, limiting the available niches due to eco-physiological factors. This study demonstrates that endemic insular deer do not evolve consistent craniometric features attributable to insular isolation, while the cranial proportions of medium-to-small-sized deer delineate a ubiquitous morphological archetype characteristic of numerous cervid taxa spanning diverse phylogenetic lineages and sharing comparable body sizes. This group of “Dama-like” deer, characterized by similar body size, metabolic rates, ecological needs, and cranial morphometry, is referred to here as the fundamental eco-physiological type, typical of warm regions within the Palearctic and Oriental biogeographic realms.
Third molars of extant-and fossil Southeast Asian deer were metrically compared using a linear-and geometric morphometric approach and discussed in relation to known taxonomic information from the literature. Our analysis suggests the presence of medium sized deer of the genus Axis and large sized taxa of the genus Cervus s. l. in Java. Axis lydekkeri and Axis javanicus are considered valid taxa, with A. lydekkeri probably related to the subgenus Hyelaphus. The large deer, such as Cervus kendengensis, Cervus stehlini and Cervus problematicus are most likely of the subgenus Rusa, the former two closely related to extant Cervus timorensis. The Sumatran fossils are members of the subgenus Rusa, but not necessarily conspecific with extant Cervus (Rusa) unicolor.
The phylogenetic pattern and timing of the radiation of Old World deer was determined based on the complete mitochondrial cytochrome b gene from 33 Cervinae taxa. Using rooted and unrooted phylogenies derived from distinct theoretical approaches, strong support was achieved for monophyly of the Old World deer with muntjacs as sister group as well as for the divergence of at least three distinct genera: Rucervus, Dama, and Cervus. The latter clade comprises what have previously been regarded as the genera or subgenera Panolia, Rusa, Cervus, Sika, and probably Przewalskium. Our data also consistently confirmed paraphyly of nominate C. elaphus and did not support the monophyly of Axis. We used these molecular phylogenies to assess the homoplastic evolution of morphological, geographical, ecological, and selected behavioural character state differences within the Cervinae. Reliable fossil calibrations, large molecular data sets, and improved dating methods are shaping a molecular time scale for the evolutionary radiation of Old World deer that occurred at the Miocene/Pliocene transition and is largely compatible with existing palaeontological evidence. Using node ages estimated from sequence data, we estimated an average per-lineage diversification rate of 0.51 ± 0.1 species per million years (my) over roughly the last 6 mya.
Zitteliana B, 2014
The report proposes a revision of poorly known fossil cervid forms and a new look on Late Neogene cervid phylogeny with implication of known already data from mitochondrial and nuclear genetic analysis. Pliocervus matheroni from Late Miocene of Europe is poorly known with unclear systematic position. Some characters (parallel orientation of long pedicles situated very close each from another; antler shape, dental morphology, developed upper canines) place P. matheroni close to Late Miocene Pavlodaria orlovi from the Hipparion fauna of Kazakhstan. Pliocervus and Pavlodaria, must be phylogenetically very close or even synonymous. Both Pliocervus and Pavlodaria share the same orientation and position on the skull of pedicles as modern Capreolus. The analysis of cranial morphology and dentition of Procapreolus moldavicus revealed a set of primitive capreoline cranial characters, combined with orientation of pedicles as in modern Odocoileus and primitive Muntiacus-like characters (large upper canines, long pedicles, deep and large preorbital fossae). P. moldavicus stands closer to a forerunner of the stock that radiated to New World deer than to modern Capreolus. Apparently, several lineages of Capreolinae evolved in Northern Eurasia before Mio-Pliocene boundary: the archaic Procapreolus lineage that does not have direct descendants in the modern fauna; the little known P. loczii that seems to be closer to the hypothesized forerunner of New World Capreolinae; and the Pliocervus-Pavlodaria lineage that may be close to modern Capreolus and Hydropotes. The dichotomy of Cervinae and Capreolinae, perhaps, is a consequence of the first expansion of Miocene ancestors of Capreolinae into the ecologically less favorable parts of Central and Boreal Asia. This may explain the generally more advanced degree of evolutionary specialization in Capreolinae, the development of additional enamel folds in the molars (the Palaeomeryx fold and the protoconal fold), which are better developed in the Capreolinae, and a more advanced specialization of premolars in the Capreolinae if compared to the Cervinae. The biogeographic expansion of Cervinae is recorded in Europe during Early Pliocene. Apparently, this expansion coincides with the first successful radiation of the Old World deer that gave such fossil forms as Arvernoceros ardei, Praeelaphus perrieri, Metacervocerus pardinensis and possibly modern Axis axis and Rucervus duvaucelii. A. ardei and R. duvaucelii share the same plane of antler construction, so Arvernoceros and Rucervus may be closely related genera, or even synonymous. The second important radiation of Cervinae took place by the end of Villafranchian, when appeared modern rich in forms Cervus, Pzewalskium, and extinct giant Praemegaceros, which together with Eucladoceros, apparently, also belongs to Rusa-Cervus stock. Dama, Megaloceros, and Megaceroides represent another stock of Cervinae that evolved in Western Eurasia and Mediterranean area.
CFS Courier Forschungsinstitut Senckenberg
A skeleton of the endemic Pleistocene Cretan deer Candiacervus sp. II (Liko, Crete, Greece) was mounted using bones of different individuals. This composite skeleton contributes to the study of the taxonomy of insular ungulates as it reveals some additional features which had not been detected in the isolated elements. Candiacervus sp. II differs from all known recent and extinct mainland deer, mainly in its proportions. Although its considerably shortened distal limbs had been already noted in the past, Candiacervus sp. II now appears at the same time to have had more or less the same vertebral column length as its large-sized continental counterparts, and a moderately upwards curved lumbar section, two features more remindful of the insular dwarf bovid Myotragus than of the small-sized mainland Axis axis. Combined with an increased massiveness of all the bones and with pronounced muscle scars, this change in body proportions indicates that Candiacervus sp. II evolved towards the n...
Paleontological Journal, 2011
The origin, trends in adaptatiogenesis and ecogenesis of megacerines reviewed. The major phylo genetic lineages are described, the main evolutionary events are dated and compared with changes in envi ronments. It is shown that the major evolutionary changes in megacerines developed against a background of considerable changes in environments, which corresponded to global climatic and great regional landscapeclimatic events.
PLoS ONE, 2011
Background: Various studies have shown that the population densities of a number of forest vertebrates, such as orangutans, are higher on Sumatra than Borneo, and that several species exhibit smaller body sizes on Borneo than Sumatra and mainland Southeast Asia. It has been suggested that differences in forest fruit productivity between the islands can explain these patterns. Here we present a large-scale comparison of forest fruit production between the islands to test this hypothesis.
Ecology and evolution, 2018
In this study we compared the phylogeographic patterns of two Rusa species, and , in order to understand what drove and maintained differentiation between these two geographically and genetically close species and investigated the route of introduction of individuals to the islands outside of the Sunda Shelf. We analyzed full mitogenomes from 56 archival samples from the distribution areas of the two species and 18 microsatellite loci in a subset of 16 individuals to generate the phylogeographic patterns of both species. Bayesian inference with fossil calibration was used to estimate the age of each species and major divergence events. Our results indicated that the split between the two species took place during the Pleistocene, ~1.8 Mya, possibly driven by adaptations of to the drier climate found on Java compared to the other islands of Sundaland. Although both markers identified two well-differentiated clades, there was a largely discrepant pattern between mitochondrial and nucl...
ZooKeys, 2016
The fluviatile terrace deposits of Khok Sung, Nakhon Ratchasima province, have yielded more than one thousand fossils, making this the richest Pleistocene vertebrate fauna of Thailand. The excellent preservation of the specimens allows precise characterization of the faunal composition. The mammalian fauna consists of fifteen species in thirteen genera, including a primate, a canid, a hyaenid, proboscideans, rhinoceroses, a suid, cervids, and bovids. Most species correspond to living taxa but globally (Stegodon cf. orientalis) and locally (Crocuta crocuta ultima, Rhinoceros unicornis, Sus barbatus, and Axis axis) extinct taxa were also present. The identification of Axis axis in Khok Sung, a chital currently restricted to the Indian Subcontinent, represents the first record of the species in Southeast Asia. Three reptilian taxa: Crocodylus cf. siamensis, Python sp., and Varanus sp., are also identified. Faunal correlations with other Southeast Asian sites suggest a late Middle to early Late Pleistocene age for the Khok Sung assemblage. However, the Khok Sung mammalian fauna is most similar to that of Thum Wiman Nakin, dated to older than 169 ka. The Khok Sung large mammal assemblage mostly comprises mainland Southeast Asian taxa that migrated to Java during the latest Middle Pleistocene, supporting the hypothesis that Thailand was a biogeographic pathway for the Sino-Malayan migration event from South China to Java.
The population of the globally endangered hog deer (Axis porcinus) has declined severely across its geographic range. Intensive monitoring of its demographic and genetic status is necessary. Northeast India is a stronghold of the species; however, in recent years the population has been getting fragmented, and it is vulnerable to extinction. We examined the demographic and genetic structure of a small hog deer population in the floating meadows of Keibul Lamjao National Park (KLNP), located on the western fringe of the Indo-Burma biodiversity hotspot for conservation planning. We used a double-observer distance sampling method to derive the hog deer abundance and population structure. We also derived the genetic diversity of the population through microsatellite screening and bottleneck detection. Our study revealed that the abundance of the deer in the park was 1.82–4.32 individuals/km2. The adult male to female ratio and fawn to doe ratio were 36.2 ± 1.9 males/100 females and 16.5...
Scientific reports, 2018
The hog deer (Axis porcinus) is threatened by habitat alteration, fragmentation, and poaching, which have led to a drastic decline of its wild population. Two subspecies of A. porcinus have been described from its distribution range. A. p. porcinus is reported to occur from Pakistan along the Himalayan foothills through Nepal, India and Myanmar, and A. p. annamiticus is found in Thailand, Indo-China, Laos, Cambodia, and Vietnam. However, the current distribution range of A. p. annamiticus is still unclear. We used the partial control region (CR) of mitochondrial DNA (mtDNA) and seven microsatellite loci to investigate the intra-species structure, differentiation, and demographic history of hog deer populations from three landscapes, the Terai Arc, Northeast, and Indo-Burma (Keibul Lamjao National Park (KLNP), Manipur, India) landscapes. We also carried out divergence time estimation using the complete mitogenome. The level of variation was ~4%, and the time of divergence of the KLNP...
PLOS ONE
The population of the globally endangered hog deer (Axis porcinus) has declined severely across its geographic range. Intensive monitoring of its demographic and genetic status is necessary. We examined the demographic and genetic structure of a small hog deer population in Keibul Lamjao National Park (KLNP), located on the western fringe of the Indo-Burma biodiversity hotspot for conservation planning. The distribution pattern of hog deer in the Park was derived based on the presence/absence of faecal pellets in 1 km × 1 km grids. We used double-observer distance sampling method to derive the hog deer abundance and population structure and compared with previous data to derive the population trend. We determined the genetic diversity of the population through microsatellite screening and bottleneck detection. The overall pellet density was 0.34 ± 0.02 pellets km-2 restricted to only 22.34 ± 0.20 km 2 area of the park. The estimated density of the deer in the park was 1.82-4.32 individuals km-2. The population showed a declining trend from 2006-08 (p < 0.05, R 2 = 0.916) with 8% annum-1 and an increasing trend from 2003-2018 (p < 0.05, R 2 = 0.9304) with 10% annum-1. The adult male-to-female ratio and fawn-to-doe ratio were 36.2 ± 1.9 males per 100 females and 16.5 ± 0.4 fawns per 100 females, respectively. The molecular examination suggested that the mean number of alleles at 23 loci was 2.70 ± 0.18, the observed heterozygosity (Ho) ranged from 0.26 to 0.63 (mean 0.42 ± 0.02), the expected heterozygosity (He) ranged from 0.23 to 0.73 (χ = 0.51 ± 0.03), and the polymorphic information content (PIC) ranged from 0.2 to 0.67 (χ = 0.43 ± 0.03) indicating a moderate level of genetic diversity. Although no bottleneck in the population was observed, the loss of genetic diversity may affect the evolutionary potential of the species at the site by limiting the selection flexibility. Conservation planning coupled with scientific management regime will help in the long term persistence of the population in the region.
Nature Ecology & Evolution, 2021
Island faunas can be characterized by gigantism in small animals and dwarfism in large animals, but the extent to which this so-called 'island rule' provides a general explanation for evolutionary trajectories on islands remains contentious. Here we use a phylogenetic metaanalysis to assess patterns and drivers of body size evolution across a global sample of paired island-mainland populations of terrestrial vertebrates. We show that 'island rule' effects are widespread in mammals, birds and reptiles, but less evident in amphibians, which mostly tend towards gigantism. We also found that the magnitude of insular dwarfism and gigantism is mediated by climate as well as island size and isolation, with more pronounced effects in smaller, more remote islands for mammals and reptiles. We conclude that the island rule is pervasive across vertebrates, but that the implications for body size evolution are nuanced and depend on an array of context-dependent ecological pressures and environmental conditions. .
Molecular Phylogenetics and Evolution, 2021
Scientific Reports
Mitochondrial DNA (mtDNA) lineages are recognized as important components of intra- and interspecific biodiversity, and allow to reveal colonization routes and phylogeographic structure of many taxa. Among these is the genus Cervus that is widely distributed across the Holarctic. We obtained sequences of complete mitochondrial genomes from 13 Cervus taxa and included them in global phylogenetic analyses of 71 Cervinae mitogenomes. The well-resolved phylogenetic trees confirmed Cervus to be monophyletic. Molecular dating based on several fossil calibration points revealed that ca. 2.6 Mya two main mitochondrial lineages of Cervus separated in Central Asia, the Western (including C. hanglu and C. elaphus) and the Eastern (comprising C. albirostris, C. canadensis and C. nippon). We also observed convergent changes in the composition of some mitochondrial genes in C. hanglu of the Western lineage and representatives of the Eastern lineage. Several subspecies of C. nippon and C. hanglu h...
Frontiers in zoology, 2018
As a small artiodactyl, the roe deer ( L.) is characterized by biological plasticity and great adaptability demonstrated by their survival under a wide variety of environmental conditions. In order to depict patterns of phenotypic variation of roe deer body this study aims to quantify variation during ontogenetic development and determine how sex-specific reproductive investment and non-uniform habitat differences relate to phenotypic variation and do these differential investments mold the patterns of phenotypic variation through modular organisation. Patterns of phenotypic correlation among body traits change during the ontogeny of roe deer, with differential influence of sex and habitat type. Modularity was found to be a feature of closed habitats with trunk+forelimbs+hindlimbs as the best supported integration/modularity hypothesis for both sexes. The indices of integration and evolvability vary with habitat type, age and sex where increased integration is followed by decreased ...
 Colin Groves
Colin Groves