CN110144384B - Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereof - Google Patents
Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereofDownload PDFInfo
- Publication number
- CN110144384B CN110144384BCN201910478044.8ACN201910478044ACN110144384BCN 110144384 BCN110144384 BCN 110144384BCN 201910478044 ACN201910478044 ACN 201910478044ACN 110144384 BCN110144384 BCN 110144384B
- Authority
- CN
- China
- Prior art keywords
- probe
- oxidative damage
- incubation
- telomeres
- chemical sensor
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108091035539telomereProteins0.000titleclaimsabstractdescription109
- 210000003411telomereAnatomy0.000titleclaimsabstractdescription107
- 102000055501telomereHuman genes0.000titleclaimsabstractdescription106
- 230000004792oxidative damageEffects0.000titleclaimsabstractdescription79
- 238000001514detection methodMethods0.000titleclaimsabstractdescription30
- 239000000126substanceSubstances0.000titleclaimsabstractdescription24
- 239000000523sampleSubstances0.000claimsabstractdescription117
- 102100033215DNA nucleotidylexotransferaseHuman genes0.000claimsabstractdescription53
- 238000007885magnetic separationMethods0.000claimsabstractdescription14
- 108010036364Deoxyribonuclease IV (Phage T4-Induced)Proteins0.000claimsabstractdescription11
- 239000006228supernatantSubstances0.000claimsabstractdescription9
- 102000004190EnzymesHuman genes0.000claimsabstractdescription7
- 108090000790EnzymesProteins0.000claimsabstractdescription7
- 230000011664signalingEffects0.000claimsabstract7
- 108020004414DNAProteins0.000claimsdescription74
- 238000000034methodMethods0.000claimsdescription51
- 238000011534incubationMethods0.000claimsdescription29
- 102000053602DNAHuman genes0.000claimsdescription27
- 230000006378damageEffects0.000claimsdescription20
- 239000011324beadSubstances0.000claimsdescription19
- 238000004557single molecule detectionMethods0.000claimsdescription15
- 108020004682Single-Stranded DNAProteins0.000claimsdescription9
- 108010090804StreptavidinProteins0.000claimsdescription9
- NKKLCOFTJVNYAQ-UHFFFAOYSA-NformamidopyrimidineChemical compoundO=CNC1=CN=CN=C1NKKLCOFTJVNYAQ-UHFFFAOYSA-N0.000claimsdescription9
- YBJHBAHKTGYVGT-ZKWXMUAHSA-N(+)-BiotinChemical compoundN1C(=O)N[C@@H]2[C@H](CCCCC(=O)O)SC[C@@H]21YBJHBAHKTGYVGT-ZKWXMUAHSA-N0.000claimsdescription6
- 239000000758substrateSubstances0.000claimsdescription6
- 108091000080PhosphotransferaseProteins0.000claimsdescription5
- CZPWVGJYEJSRLH-UHFFFAOYSA-NPyrimidineChemical compoundC1=CN=CN=C1CZPWVGJYEJSRLH-UHFFFAOYSA-N0.000claimsdescription5
- ARLKCWCREKRROD-POYBYMJQSA-N[[(2s,5r)-5-(4-amino-2-oxopyrimidin-1-yl)oxolan-2-yl]methoxy-hydroxyphosphoryl] phosphono hydrogen phosphateChemical compoundO=C1N=C(N)C=CN1[C@@H]1O[C@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)CC1ARLKCWCREKRROD-POYBYMJQSA-N0.000claimsdescription5
- 229960002685biotinDrugs0.000claimsdescription5
- 239000011616biotinSubstances0.000claimsdescription5
- 102000020233phosphotransferaseHuman genes0.000claimsdescription5
- 239000002157polynucleotideSubstances0.000claimsdescription5
- 239000007795chemical reaction productSubstances0.000claimsdescription4
- 238000002189fluorescence spectrumMethods0.000claimsdescription4
- 235000020958biotinNutrition0.000claimsdescription3
- 201000010099diseaseDiseases0.000claimsdescription3
- 208000037265diseases, disorders, signs and symptomsDiseases0.000claimsdescription3
- 230000005284excitationEffects0.000claimsdescription3
- 239000012634fragmentSubstances0.000claimsdescription3
- 230000004048modificationEffects0.000claimsdescription2
- 238000012986modificationMethods0.000claimsdescription2
- 238000003745diagnosisMethods0.000claims2
- 230000006287biotinylationEffects0.000claims1
- 238000007413biotinylationMethods0.000claims1
- 230000027832depurinationEffects0.000claims1
- 239000002245particleSubstances0.000claims1
- 108010008286DNA nucleotidylexotransferaseProteins0.000abstractdescription49
- 238000003776cleavage reactionMethods0.000abstractdescription19
- 230000007017scissionEffects0.000abstractdescription19
- 230000001404mediated effectEffects0.000abstractdescription13
- 238000006116polymerization reactionMethods0.000abstractdescription12
- 125000004122cyclic groupChemical group0.000abstractdescription9
- 238000003384imaging methodMethods0.000abstractdescription9
- 230000008439repair processEffects0.000abstractdescription4
- 210000004027cellAnatomy0.000description32
- 230000035945sensitivityEffects0.000description14
- 239000000047productSubstances0.000description12
- 230000015572biosynthetic processEffects0.000description9
- 230000001590oxidative effectEffects0.000description9
- 238000003786synthesis reactionMethods0.000description9
- 238000006243chemical reactionMethods0.000description8
- 238000002474experimental methodMethods0.000description8
- UYTPUPDQBNUYGX-UHFFFAOYSA-NguanineChemical compoundO=C1NC(N)=NC2=C1N=CN2UYTPUPDQBNUYGX-UHFFFAOYSA-N0.000description8
- 239000011535reaction bufferSubstances0.000description8
- 108010021757Polynucleotide 5'-Hydroxyl-KinaseProteins0.000description7
- 102000008422Polynucleotide 5'-hydroxyl-kinaseHuman genes0.000description7
- 238000010586diagramMethods0.000description7
- 230000003321amplificationEffects0.000description6
- 238000003199nucleic acid amplification methodMethods0.000description6
- 230000008789oxidative DNA damageEffects0.000description6
- 239000000243solutionSubstances0.000description6
- 238000005820transferase reactionMethods0.000description6
- 108091028043Nucleic acid sequenceProteins0.000description5
- 238000004458analytical methodMethods0.000description5
- 230000029087digestionEffects0.000description5
- 238000004128high performance liquid chromatographyMethods0.000description5
- 239000000203mixtureSubstances0.000description5
- 239000002773nucleotideSubstances0.000description5
- 125000003729nucleotide groupChemical group0.000description5
- 108010000577DNA-Formamidopyrimidine GlycosylaseProteins0.000description4
- 206010028980NeoplasmDiseases0.000description4
- FAPWRFPIFSIZLT-UHFFFAOYSA-MSodium chlorideChemical compound[Na+].[Cl-]FAPWRFPIFSIZLT-UHFFFAOYSA-M0.000description4
- 230000008901benefitEffects0.000description4
- 239000000872bufferSubstances0.000description4
- 201000011510cancerDiseases0.000description4
- SUYVUBYJARFZHO-RRKCRQDMSA-NdATPChemical compoundC1=NC=2C(N)=NC=NC=2N1[C@H]1C[C@H](O)[C@@H](COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1SUYVUBYJARFZHO-RRKCRQDMSA-N0.000description4
- SUYVUBYJARFZHO-UHFFFAOYSA-NdATPNatural productsC1=NC=2C(N)=NC=NC=2N1C1CC(O)C(COP(O)(=O)OP(O)(=O)OP(O)(O)=O)O1SUYVUBYJARFZHO-UHFFFAOYSA-N0.000description4
- 238000001502gel electrophoresisMethods0.000description4
- QKNYBSVHEMOAJP-UHFFFAOYSA-N2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chlorideChemical compoundCl.OCC(N)(CO)COQKNYBSVHEMOAJP-UHFFFAOYSA-N0.000description3
- KCXVZYZYPLLWCC-UHFFFAOYSA-NEDTAChemical compoundOC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=OKCXVZYZYPLLWCC-UHFFFAOYSA-N0.000description3
- MHAJPDPJQMAIIY-UHFFFAOYSA-NHydrogen peroxideChemical compoundOOMHAJPDPJQMAIIY-UHFFFAOYSA-N0.000description3
- HEMHJVSKTPXQMS-UHFFFAOYSA-MSodium hydroxideChemical compound[OH-].[Na+]HEMHJVSKTPXQMS-UHFFFAOYSA-M0.000description3
- 230000033590base-excision repairEffects0.000description3
- 238000005516engineering processMethods0.000description3
- 210000005260human cellAnatomy0.000description3
- 230000003902lesionEffects0.000description3
- UEZVMMHDMIWARA-UHFFFAOYSA-MphosphonateChemical group[O-]P(=O)=OUEZVMMHDMIWARA-UHFFFAOYSA-M0.000description3
- 238000002264polyacrylamide gel electrophoresisMethods0.000description3
- 239000011541reaction mixtureSubstances0.000description3
- 230000003252repetitive effectEffects0.000description3
- 230000004044responseEffects0.000description3
- 235000011178triphosphateNutrition0.000description3
- 239000001226triphosphateSubstances0.000description3
- 125000002264triphosphate groupChemical class[H]OP(=O)(O[H])OP(=O)(O[H])OP(=O)(O[H])O*0.000description3
- KDELTXNPUXUBMU-UHFFFAOYSA-N2-[2-[bis(carboxymethyl)amino]ethyl-(carboxymethyl)amino]acetic acid boric acidChemical compoundOB(O)O.OB(O)O.OB(O)O.OC(=O)CN(CC(O)=O)CCN(CC(O)=O)CC(O)=OKDELTXNPUXUBMU-UHFFFAOYSA-N0.000description2
- 108020001738DNA GlycosylaseProteins0.000description2
- 102000028381DNA glycosylaseHuman genes0.000description2
- 239000006144Dulbecco’s modified Eagle's mediumSubstances0.000description2
- 230000000694effectsEffects0.000description2
- 238000000835electrochemical detectionMethods0.000description2
- 239000012091fetal bovine serumSubstances0.000description2
- 238000002290gas chromatography-mass spectrometryMethods0.000description2
- 231100000025genetic toxicologyToxicity0.000description2
- 230000001738genotoxic effectEffects0.000description2
- 238000000589high-performance liquid chromatography-mass spectrometryMethods0.000description2
- 239000013642negative controlSubstances0.000description2
- 230000036542oxidative stressEffects0.000description2
- 108090000623proteins and genesProteins0.000description2
- 238000004445quantitative analysisMethods0.000description2
- 239000003642reactive oxygen metaboliteSubstances0.000description2
- 238000003753real-time PCRMethods0.000description2
- 108091008146restriction endonucleasesProteins0.000description2
- 239000011780sodium chlorideSubstances0.000description2
- 238000012360testing methodMethods0.000description2
- YKBGVTZYEHREMT-KVQBGUIXSA-N2'-deoxyguanosineChemical compoundC1=NC=2C(=O)NC(N)=NC=2N1[C@H]1C[C@H](O)[C@@H](CO)O1YKBGVTZYEHREMT-KVQBGUIXSA-N0.000description1
- ZKHQWZAMYRWXGA-KQYNXXCUSA-JATP(4-)Chemical compoundC1=NC=2C(N)=NC=NC=2N1[C@@H]1O[C@H](COP([O-])(=O)OP([O-])(=O)OP([O-])([O-])=O)[C@@H](O)[C@H]1OZKHQWZAMYRWXGA-KQYNXXCUSA-J0.000description1
- 208000010507Adenocarcinoma of LungDiseases0.000description1
- ZKHQWZAMYRWXGA-UHFFFAOYSA-NAdenosine triphosphateNatural productsC1=NC=2C(N)=NC=NC=2N1C1OC(COP(O)(=O)OP(O)(=O)OP(O)(O)=O)C(O)C1OZKHQWZAMYRWXGA-UHFFFAOYSA-N0.000description1
- 108091023037AptamerProteins0.000description1
- 108091003079Bovine Serum AlbuminProteins0.000description1
- 208000024172Cardiovascular diseaseDiseases0.000description1
- 206010008342Cervix carcinomaDiseases0.000description1
- 206010009944Colon cancerDiseases0.000description1
- 238000007399DNA isolationMethods0.000description1
- 102000010719DNA-(Apurinic or Apyrimidinic Site) LyaseHuman genes0.000description1
- 108010063362DNA-(Apurinic or Apyrimidinic Site) LyaseProteins0.000description1
- 241000588724Escherichia coliSpecies0.000description1
- 208000031448Genomic InstabilityDiseases0.000description1
- 101001121408Homo sapiens L-amino-acid oxidaseProteins0.000description1
- 102100026388L-amino-acid oxidaseHuman genes0.000description1
- 229910019142PO4Inorganic materials0.000description1
- 238000011529RT qPCRMethods0.000description1
- 108091081062Repeated sequence (DNA)Proteins0.000description1
- 239000007984Tris EDTA bufferSubstances0.000description1
- 208000006105Uterine Cervical NeoplasmsDiseases0.000description1
- 238000009825accumulationMethods0.000description1
- 150000003838adenosinesChemical class0.000description1
- 238000003556assayMethods0.000description1
- 230000009286beneficial effectEffects0.000description1
- 230000031018biological processes and functionsEffects0.000description1
- KGBXLFKZBHKPEV-UHFFFAOYSA-Nboric acidChemical compoundOB(O)OKGBXLFKZBHKPEV-UHFFFAOYSA-N0.000description1
- 239000004327boric acidSubstances0.000description1
- 238000005251capillar electrophoresisMethods0.000description1
- 238000010523cascade reactionMethods0.000description1
- 230000003197catalytic effectEffects0.000description1
- 238000004113cell cultureMethods0.000description1
- 230000003833cell viabilityEffects0.000description1
- 201000010881cervical cancerDiseases0.000description1
- 210000000349chromosomeAnatomy0.000description1
- 208000029742colonic neoplasmDiseases0.000description1
- 230000021615conjugationEffects0.000description1
- 238000007796conventional methodMethods0.000description1
- 230000008878couplingEffects0.000description1
- 238000010168coupling processMethods0.000description1
- 238000005859coupling reactionMethods0.000description1
- 238000007405data analysisMethods0.000description1
- 230000007812deficiencyEffects0.000description1
- 206010012601diabetes mellitusDiseases0.000description1
- 238000007865dilutingMethods0.000description1
- 238000010790dilutionMethods0.000description1
- 239000012895dilutionSubstances0.000description1
- 238000010494dissociation reactionMethods0.000description1
- 230000005593dissociationsEffects0.000description1
- 239000003814drugSubstances0.000description1
- 238000013399early diagnosisMethods0.000description1
- 230000004049epigenetic modificationEffects0.000description1
- 238000002795fluorescence methodMethods0.000description1
- 238000002073fluorescence micrographMethods0.000description1
- PCHJSUWPFVWCPO-UHFFFAOYSA-NgoldChemical compound[Au]PCHJSUWPFVWCPO-UHFFFAOYSA-N0.000description1
- 239000010931goldSubstances0.000description1
- 229910052737goldInorganic materials0.000description1
- 125000002887hydroxy groupChemical group[H]O*0.000description1
- 238000007654immersionMethods0.000description1
- 238000001727in vivoMethods0.000description1
- 230000000977initiatory effectEffects0.000description1
- 238000012417linear regressionMethods0.000description1
- 210000005229liver cellAnatomy0.000description1
- 201000005249lung adenocarcinomaDiseases0.000description1
- 238000005259measurementMethods0.000description1
- 230000007246mechanismEffects0.000description1
- 238000005457optimizationMethods0.000description1
- 239000007800oxidant agentSubstances0.000description1
- 230000003647oxidationEffects0.000description1
- 238000007254oxidation reactionMethods0.000description1
- 239000010452phosphateSubstances0.000description1
- 238000002360preparation methodMethods0.000description1
- 230000008569processEffects0.000description1
- 230000009145protein modificationEffects0.000description1
- 230000010076replicationEffects0.000description1
- 238000000492total internal reflection fluorescence microscopyMethods0.000description1
- 230000002103transcriptional effectEffects0.000description1
- 238000010200validation analysisMethods0.000description1
Images








Classifications
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12Q—MEASURING OR TESTING PROCESSES INVOLVING ENZYMES, NUCLEIC ACIDS OR MICROORGANISMS; COMPOSITIONS OR TEST PAPERS THEREFOR; PROCESSES OF PREPARING SUCH COMPOSITIONS; CONDITION-RESPONSIVE CONTROL IN MICROBIOLOGICAL OR ENZYMOLOGICAL PROCESSES
- C12Q1/00—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions
- C12Q1/68—Measuring or testing processes involving enzymes, nucleic acids or microorganisms; Compositions therefor; Processes of preparing such compositions involving nucleic acids
- C12Q1/6813—Hybridisation assays
- C12Q1/6816—Hybridisation assays characterised by the detection means
- C12Q1/6825—Nucleic acid detection involving sensors
Landscapes
- Chemical & Material Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Organic Chemistry (AREA)
- Analytical Chemistry (AREA)
- Zoology (AREA)
- Wood Science & Technology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Health & Medical Sciences (AREA)
- Engineering & Computer Science (AREA)
- Microbiology (AREA)
- Immunology (AREA)
- Molecular Biology (AREA)
- Biotechnology (AREA)
- Biophysics (AREA)
- Physics & Mathematics (AREA)
- Biochemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Genetics & Genomics (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
Translated fromChinese技术领域Technical Field
本发明属于生物检测和分子生物学技术领域,具体涉及一种检测端粒内氧化性损伤的荧光化学传感器及其检测方法和应用。The invention belongs to the technical field of biological detection and molecular biology, and specifically relates to a fluorescent chemical sensor for detecting oxidative damage in telomeres and a detection method and application thereof.
背景技术Background Art
公开该背景技术部分的信息仅仅旨在增加对本发明的总体背景的理解,而不必然被视为承认或以任何形式暗示该信息构成已经成为本领域一般技术人员所公知的现有技术。The information disclosed in this background technology section is only intended to enhance the understanding of the overall background of the invention, and should not necessarily be regarded as an admission or any form of suggestion that the information constitutes the prior art already known to a person skilled in the art.
端粒由双链的重复序列(TTAGGG)n组成且具有单链的突出末端。端粒封闭真核染色体的末端并在基因组的稳定性和细胞活力中发挥关键作用。TTAGGG重复序列的富含G的特性使端粒极易受氧化损伤,特别是在单链区域,因为它更容易与氧化剂反应。氧化性损伤8-oxoG和FapyG是脱氧鸟苷(dG)最常见的氧化产物,可以通过引起复制错误和转录干扰诱导遗传毒性。端粒中富含的G碱基有助于氧化应激过程中G氧化性损伤的优先积累,而且它的修复效率比基因组中其他序列中的G损伤偏低。端粒的氧化损伤与端粒长度和完整性的变化有关,这将导致多种疾病的发生,如心血管疾病,糖尿病和癌症。因此,检测端粒中的氧化损伤水平将为分子医学,早期诊断和化学品的遗传毒性提供巨大益处。Telomeres consist of double-stranded repeat sequences (TTAGGG)n with single-stranded protruding ends. Telomeres cap the ends of eukaryotic chromosomes and play a key role in genome stability and cell viability. The G-rich nature of the TTAGGG repeat sequence makes telomeres extremely susceptible to oxidative damage, especially in the single-stranded region, because it reacts more easily with oxidants. Oxidative damage 8-oxoG and FapyG are the most common oxidation products of deoxyguanosine (dG), which can induce genotoxicity by causing replication errors and transcriptional interference. The rich G bases in telomeres contribute to the preferential accumulation of G oxidative damage during oxidative stress, and its repair efficiency is lower than that of G damage in other sequences in the genome. Oxidative damage to telomeres is associated with changes in telomere length and integrity, which will lead to the occurrence of multiple diseases, such as cardiovascular disease, diabetes and cancer. Therefore, detecting the level of oxidative damage in telomeres will provide great benefits for molecular medicine, early diagnosis and genotoxicity of chemicals.
目前,检测基因组中8-oxoG的最常用方法包括高效液相色谱(HPLC)与电化学检测相结合(HPLC-ECD)以及HPLC与气相色谱-质谱联用(HPLC-MS)/MS)。这些方法可以提供更准确的8-oxoG计数,但需要把DNA序列消化成单个核苷酸碱基,而且需要复杂的仪器设备及繁琐的实验步骤。迄今为止,由于低丰度和高度重复的基因区域,很少有方法可用于检测端粒中的氧化性损伤水平。最近,纳米孔技术和基于定量PCR(q-PCR)的方法可以用于检测人端粒序列中的氧化性损伤。然而,发明人发现,它们需要特定的损伤标记和复杂的热循环仪来精确控制热循环。利用腺苷的衍生物三磷酸Adap可用于区分人端粒序列中的8-oxoG损伤和正常的G碱基,但是该方法难以定量8-oxoG损伤。Currently, the most commonly used methods for detecting 8-oxoG in the genome include high performance liquid chromatography (HPLC) combined with electrochemical detection (HPLC-ECD) and HPLC combined with gas chromatography-mass spectrometry (HPLC-MS/MS). These methods can provide more accurate 8-oxoG counts, but require digestion of DNA sequences into single nucleotide bases, and require complex instrumentation and cumbersome experimental steps. To date, due to the low abundance and highly repetitive gene regions, few methods have been available to detect the level of oxidative damage in telomeres. Recently, nanopore technology and quantitative PCR (q-PCR)-based methods can be used to detect oxidative damage in human telomere sequences. However, the inventors found that they require specific damage markers and complex thermal cyclers to accurately control thermal cycles. The use of adenosine triphosphate Adap, a derivative of adenosine, can be used to distinguish 8-oxoG damage from normal G bases in human telomere sequences, but this method is difficult to quantify 8-oxoG damage.
发明内容Summary of the invention
针对上述现有技术的不足,本发明提供一种检测端粒内氧化性损伤的荧光化学传感器及其检测方法和应用。本发明中将修复酶的特异性与末端脱氧核苷酸转移酶(TdT)介导的聚合延伸相结合,以启动核酸内切酶IV(Endo IV)诱导的循环切割信号探针,诱导EndoIV裂解信号探针中的AP位点,导致信号探针上AF488荧光团的释放,同时产生具有游离3'-OH末端的信号探针。具有游离3'-OH的每个信号探针可以通过TdT有效地延长以产生长的poly-A序列。释放的poly-A序列和新产生的poly-A序列可以诱导信号探针的多次循环切割,释放出更多的AF488荧光团。在磁力分离下,上清液中的荧光团可以通过单分子成像定量。本发明无需复杂的仪器或繁琐的程序,同时检测灵敏度高,具有良好的实际应用之价值。In view of the deficiencies of the above-mentioned prior art, the present invention provides a fluorescent chemical sensor for detecting oxidative damage in telomeres, and a detection method and application thereof. In the present invention, the specificity of the repair enzyme is combined with the terminal deoxynucleotidyl transferase (TdT)-mediated polymerization extension to initiate the cyclic cutting of the signal probe induced by endonuclease IV (Endo IV), induce Endo IV to cleave the AP site in the signal probe, resulting in the release of the AF488 fluorophore on the signal probe, and simultaneously generate a signal probe with a free 3'-OH end. Each signal probe with a free 3'-OH can be effectively extended by TdT to generate a long poly-A sequence. The released poly-A sequence and the newly generated poly-A sequence can induce multiple cyclic cutting of the signal probe to release more AF488 fluorophores. Under magnetic separation, the fluorophore in the supernatant can be quantified by single-molecule imaging. The present invention does not require complex instruments or cumbersome procedures, and has high detection sensitivity and good practical application value.
本发明是通过如下技术方案实现的:The present invention is achieved through the following technical solutions:
本发明的第一个方面,提供一种检测端粒内氧化性损伤的荧光化学传感器,所述荧光化学传感器至少包括信号探针、捕获探针和辅助探针。In a first aspect, the present invention provides a fluorescent chemical sensor for detecting oxidative damage in telomeres, wherein the fluorescent chemical sensor comprises at least a signal probe, a capture probe and an auxiliary probe.
其中,所述信号探针为具有脱嘌呤嘧啶位点(AP位点)的富T单链DNA序列,所述信号探针5'端修饰有生物素,所述信号探针3'端修饰有荧光基团,优选使用AF488进行修饰;Wherein, the signal probe is a T-rich single-stranded DNA sequence with an apurinic pyrimidine site (AP site), the 5' end of the signal probe is modified with biotin, and the 3' end of the signal probe is modified with a fluorescent group, preferably AF488;
进一步的,所述信号探针与链霉抗生物素蛋白包被的磁珠进行缀合。Furthermore, the signal probe is conjugated to magnetic beads coated with streptavidin.
所述捕获探针用于捕获富集端粒,其具有与端粒杂交的单链DNA序列,所述捕获探针3'端进行生物素化修饰;The capture probe is used to capture and enrich telomeres, and has a single-stranded DNA sequence that hybridizes with the telomeres. The 3' end of the capture probe is biotinylated;
所述辅助探针为3'端用ddC修饰的单链DNA序列,其与单链端粒杂交形成甲酰胺嘧啶[fapy]-DNA糖基化酶的dsDNA底物。The auxiliary probe is a single-stranded DNA sequence modified with ddC at the 3' end, which hybridizes with the single-stranded telomere to form a dsDNA substrate for formamidopyrimidine [fapy]-DNA glycosylase.
进一步的,所述荧光化学传感器还包括甲酰胺嘧啶[fapy]-DNA糖基化酶、末端脱氧核苷酸转移酶(TdT)、核酸内切酶IV和T4-多核苷酸激酶。Furthermore, the fluorescent chemical sensor also includes formamidopyrimidine [fapy]-DNA glycosylase, terminal deoxynucleotidyl transferase (TdT), endonuclease IV and T4-polynucleotide kinase.
进一步的,所述端粒内氧化性损伤包括8-oxoG损伤。Furthermore, the oxidative damage in the telomere includes 8-oxoG damage.
本发明的第二个方面,提供基于上述荧光化学传感器检测端粒内氧化性损伤的方法,所述方法包括:The second aspect of the present invention provides a method for detecting oxidative damage in telomeres based on the above-mentioned fluorescent chemical sensor, the method comprising:
S1、用dsDNA片段酶消化提取基因组DNA,获得具有50-200bp的DNA;S1. Extract genomic DNA by digestion with dsDNA fragmentase to obtain DNA with 50-200 bp;
S2、加入捕获探针和链霉抗生物素蛋白包被的磁珠孵育处理,捕获探针用于捕获富集端粒;S2, adding capture probes and streptavidin-coated magnetic beads for incubation, where the capture probes are used to capture and enrich telomeres;
S3、加入TdT进行孵育处理,催化三磷酸脱氧核苷酸(dATPs)重复加入DNA分子的3'-羟基末端(3'-OH);S3, adding TdT for incubation treatment, catalyzing the repeated addition of deoxynucleotide triphosphates (dATPs) to the 3'-hydroxyl end (3'-OH) of the DNA molecule;
S4、加入辅助探针、甲酰胺嘧啶[fapy]-DNA糖基化酶进行孵育处理,其中辅助探针用于与单链端粒杂交以形成甲酰胺嘧啶[fapy]-DNA糖基化酶的dsDNA底物;S4, adding an auxiliary probe and fapy-DNA glycosylase for incubation, wherein the auxiliary probe is used to hybridize with the single-stranded telomere to form a dsDNA substrate for fapy-DNA glycosylase;
S5、加入TdT、核酸内切酶IV和磁珠偶联信号探针进行孵育处理;S5, adding TdT, endonuclease IV and magnetic bead-coupled signal probe for incubation;
S6、磁分离后,对上清液进行单分子检测。S6. After magnetic separation, the supernatant is subjected to single molecule detection.
本发明的第三个方面,提供上述荧光化学传感器和/或检测方法在检测端粒内氧化性损伤中的应用。所述端粒内氧化性损伤包括8-oxoG损伤。The third aspect of the present invention provides the use of the fluorescent chemical sensor and/or detection method in detecting oxidative damage in telomeres. The oxidative damage in telomeres includes 8-oxoG damage.
本发明首次提出了能够检测和量化人类端粒氧化损伤的超灵敏单分子成像技术。单分子检测是生物分子分析的有力工具,具有灵敏度高,样品消耗低,信噪比高的优点,并已成为DNA糖基化酶,DNA表观遗传修饰,蛋白质修饰和生物过程动态研究的有前途的方法。This invention presents for the first time an ultrasensitive single-molecule imaging technique capable of detecting and quantifying oxidative damage to human telomeres. Single-molecule detection is a powerful tool for biomolecular analysis with the advantages of high sensitivity, low sample consumption, and high signal-to-noise ratio, and has become a promising method for the study of DNA glycosylases, DNA epigenetic modifications, protein modifications, and dynamics of biological processes.
在本发明中,将修复酶的特异性与末端脱氧核苷酸转移酶(TdT)介导的聚合延伸相结合,以启动核酸内切酶IV(Endo IV)诱导的循环切割信号探针,导致AF488荧光团的释放。值得注意的是,TdT是一种无模板的聚合酶,它可以催化三磷酸脱氧核苷酸(dATPs)重复加入DNA分子的3'-羟基末端(3'-OH)。因此,每个端粒片段在氧化受损碱基的位置产生3'-OH,并且通过TdT有效地延长产生长的poly-A序列。信号探针与生成的polyA序列结合之后,诱导Endo IV裂解信号探针中的AP位点,释放出AF488荧光团,poly-A序列,同时产生具有游离3'-OH末端的的信号探针。具有游离3'-OH的每个信号探针可以通过TdT有效地延长以产生长的poly-A序列。释放的poly-A序列和新产生的poly-A序列可以诱导信号探针的多次循环切割,释放出更多的AF488荧光团。在磁力分离下,上清液中的荧光团可以通过单分子成像定量。该方法可以在混合体系中检测到0.001%的氧化性损伤。此外,该方法可以灵敏地检测不同浓度的H2O2诱导的氧化应激中端粒内氧化性损伤的变化,甚至可以检测到从1000μM H2O2处理的HeLa细胞中提取的0.1ng基因组DNA中端粒内的氧化性损伤。而且通过标准的线性关系方程可以计算出每个HeLa细胞中端粒中含有34-44个氧化性损伤位点。In the present invention, the specificity of the repair enzyme is combined with terminal deoxynucleotidyl transferase (TdT)-mediated polymerization extension to initiate endonuclease IV (Endo IV)-induced cyclic cleavage of the signal probe, resulting in the release of the AF488 fluorophore. It is noteworthy that TdT is a template-free polymerase that can catalyze the repeated addition of deoxynucleotide triphosphates (dATPs) to the 3'-hydroxyl end (3'-OH) of the DNA molecule. Therefore, each telomere fragment produces a 3'-OH at the position of the oxidized damaged base, and is effectively extended by TdT to produce a long poly-A sequence. After the signal probe binds to the generated polyA sequence, Endo IV is induced to cleave the AP site in the signal probe, releasing the AF488 fluorophore, the poly-A sequence, and simultaneously generating a signal probe with a free 3'-OH end. Each signal probe with a free 3'-OH can be effectively extended by TdT to produce a long poly-A sequence. The released poly-A sequence and the newly generated poly-A sequence can induce multiple cyclic cleavages of the signal probe, releasing more AF488 fluorophores. Under magnetic separation, the fluorophores in the supernatant can be quantified by single-molecule imaging. This method can detect 0.001% oxidative damage in a mixed system. In addition, this method can sensitively detect changes in oxidative damage in telomeres during oxidative stress induced by different concentrations of H2O2, and can even detect oxidative damage in telomeres in 0.1ng genomic DNA extracted from HeLa cells treated with 1000μM H2O2. And the standard linear relationship equation can be used to calculate that each HeLa cell telomere contains 34-44 oxidative damage sites.
本发明有益效果:Beneficial effects of the present invention:
(1)本发明整个反应在等温条件下进行:传统的检测8-oxoG的方法往往需要复杂的仪器或繁琐的程序,与之相比,本发明整个反应在等温条件下进行不需要复杂的仪器或繁琐的程序。(1) The entire reaction of the present invention is carried out under isothermal conditions: Traditional methods for detecting 8-oxoG often require complex instruments or cumbersome procedures. In contrast, the entire reaction of the present invention is carried out under isothermal conditions without the need for complex instruments or cumbersome procedures.
(2)本发明利用TDT和Endo IV的结合不仅消除了模板或特异性限制酶识别序列对切口酶切裂的参与还大大提高了检测灵敏度。TDT介导的模板非依赖性聚合延伸和Endo IV介导的信号探针切割消除了模板或特异性限制酶识别序列对切口酶切裂的参与,并且TDT和Endo IV的结合可诱导信号探针的环状切割和延伸,触发后续的信号放大级联反应,大大提高了检测灵敏度。(2) The present invention utilizes the combination of TDT and Endo IV to not only eliminate the participation of the template or specific restriction enzyme recognition sequence in the nicking enzyme cleavage, but also greatly improve the detection sensitivity. TDT-mediated template-independent polymerization extension and Endo IV-mediated signal probe cleavage eliminate the participation of the template or specific restriction enzyme recognition sequence in the nicking enzyme cleavage, and the combination of TDT and Endo IV can induce the circular cleavage and extension of the signal probe, triggering the subsequent signal amplification cascade reaction, greatly improving the detection sensitivity.
(3)本发明引入磁分离和单分子法的高信噪比提供了接近零的背景信号,进一步提高了检测灵敏度。单分子检测是生物分子分析的有力工具,具有灵敏度高,样品消耗低,信噪比高的优点。本发明引入单分子法的高信噪比使得背景信号接近于零,进一步提高了检测的灵敏度。利用低背景信号,结合高效TdT激活的Endo IV催化信号放大和单分子检测的高灵敏度,该方法对氧化鸟嘌呤损伤具有超高灵敏度,检出限低至9.3×10-17M,它可以识别低丰度的氧化鸟嘌呤损伤的存在,低至0.001%。具有良好的实际应用之前景。(3) The high signal-to-noise ratio of magnetic separation and single-molecule method introduced in the present invention provides a background signal close to zero, further improving the detection sensitivity. Single-molecule detection is a powerful tool for biomolecular analysis, with the advantages of high sensitivity, low sample consumption, and high signal-to-noise ratio. The high signal-to-noise ratio of the single-molecule method introduced in the present invention makes the background signal close to zero, further improving the sensitivity of detection. Utilizing low background signals, combined with efficient TdT-activated Endo IV catalytic signal amplification and high sensitivity of single-molecule detection, this method has ultra-high sensitivity to oxidative guanine damage, with a detection limit as low as 9.3×10-17 M, and it can identify the presence of low-abundance oxidative guanine damage, as low as 0.001%. It has good prospects for practical application.
附图说明BRIEF DESCRIPTION OF THE DRAWINGS
构成本发明的一部分的说明书附图用来提供对本发明的进一步理解,本发明的示意性实施例及其说明用于解释本发明,并不构成对本发明的不当限定。The accompanying drawings in the specification, which constitute a part of the present invention, are used to provide a further understanding of the present invention. The exemplary embodiments of the present invention and their descriptions are used to explain the present invention and do not constitute improper limitations on the present invention.
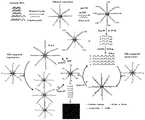
图1为本发明实施例1中荧光化学传感器检测端粒内氧化性损伤的机理图。FIG1 is a diagram showing the mechanism of the fluorescent chemical sensor in Example 1 of the present invention for detecting oxidative damage in telomeres.
图2为检测可行性的实验图;图2A为本发明实施例1中TdT介导的去除8-oxoG碱基后聚合反应的凝胶电泳成像图;泳道1,0.5μMTero-1/辅助探针+3U PNK+4U TdT;泳道2,0.5μMTero-1/辅助探针+1.6U Fpg+3U PNK+4U TdT;图2B为本发明实施例1中含有两个8-oxoG碱基的10nM(Telo-2)和10nM正常端粒(Telo-0)的荧光信号图;图2C为检测10pM Telo-2(I)和10pM Telo-0(II)的单分子成像得到的的AF488的荧光图;比例尺为2.5μm。Figure 2 is an experimental diagram of the detection feasibility; Figure 2A is a gel electrophoresis imaging diagram of the polymerization reaction after TdT-mediated removal of 8-oxoG bases in Example 1 of the present invention;
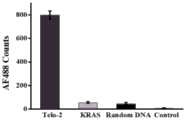
图3为分别检测含有两个8-oxoG碱基的1nM端粒(Telo-2),1nM含有两个8-oxoG碱基的合成KRAS序列(KRAS),1nM含有两个8-oxoG碱基随机的合成DNA序列(Random DNA)和反应缓冲液(control)的AF488计数图。误差棒为三组实验的标准偏差。Figure 3 is a graph of AF488 counts for detecting 1 nM telomere containing two 8-oxoG bases (Telo-2), 1 nM synthetic KRAS sequence containing two 8-oxoG bases (KRAS), 1 nM random synthetic DNA sequence containing two 8-oxoG bases (Random DNA) and reaction buffer (control). The error bars are the standard deviations of three sets of experiments.
图4为在最佳实验条件下,氧化损伤检测方法的灵敏度示意图;图4A为在1×10-16至1×10-11M范围内的氧化碱基损伤浓度对应的AF488的对数线性相关性图;图4B为在Telo-2和Telo-0的混合物中测出与实际输入氧化损伤水平之间的相关性图;其中,Telo-2和Telo-0的总浓度为10pM。误差棒为三组实验的标准偏差。Figure 4 is a schematic diagram of the sensitivity of the oxidative damage detection method under optimal experimental conditions; Figure 4A is a log-linear correlation diagram of AF488 corresponding to the oxidative base damage concentration in the range of 1×10-16 to 1×10-11 M; Figure 4B is a correlation diagram between the measured and actual input oxidative damage levels in a mixture of Telo-2 and Telo-0; where the total concentration of Telo-2 and Telo-0 is 10pM. Error bars are standard deviations of three sets of experiments.
图5为由活性氧引起的基因组DNA端粒内的氧化性损伤检测图;图5A为暴露于不同浓度的H2O2的HeLa细胞的端粒中的氧化损伤的AF488计数变化图;其中,端粒是从50ng基因组DNA中分离出;图5B为在0.1至100ng范围内,AF488计数与来自1000μMH2O2处理的HeLa细胞的基因组DNA量的对数线性关系图。误差棒为三组实验的标准偏差。Figure 5 is a graph showing the detection of oxidative damage in genomic DNA telomeres caused by reactive oxygen species; Figure 5A is a graph showing the changes in AF488 counts of oxidative damage in telomeres of HeLa cells exposed to different concentrations of H2 O2 ; wherein telomeres were isolated from 50 ng of genomic DNA; Figure 5B is a graph showing the logarithmic linear relationship between AF488 counts and the amount of genomic DNA from HeLa cells treated with 1000 μMH 2 O2 in the range of 0.1 to 100 ng. Error bars are standard deviations of three sets of experiments.
图6为Fpg诱导的氧化损伤碱基的去除的凝胶电泳成像图;其中,泳道1,0.5μMTero-Cy5/辅助探针;泳道2,0.5μM Telo-Cy5/辅助探针+1.6U Fpg。FIG6 is a gel electrophoresis image of the removal of oxidatively damaged bases induced by Fpg; wherein,
图7为Endo IV的量对应的AF488计数图。Telo-2浓度为10pM。误差棒为三组实验的标准偏差。Figure 7 is a graph showing the AF488 counts corresponding to the amount of Endo IV. The Telo-2 concentration was 10 pM. The error bars are the standard deviation of three sets of experiments.
图8为TdT的量对应的AF488计数图;其中,Telo-2浓度为10pM。误差棒为三组实验的标准偏差。Figure 8 is a graph showing the AF488 counts corresponding to the amount of TdT, wherein the Telo-2 concentration is 10 pM. The error bars are the standard deviations of three sets of experiments.
图9为输入的氧化损伤在含有两个8-oxoG碱基(Telo-2)和没有氧化碱基损伤(Telo-0)的正常端粒的混合物的AF488计数图。误差棒为三组实验的标准偏差。Figure 9 is a graph of AF488 counts of input oxidative damage in a mixture of normal telomeres containing two 8-oxoG bases (Telo-2) and no oxidative base damage (Telo-0). Error bars are standard deviations of three experiments.
图10为来自用1000μMH2O2处理的不同人细胞系(即,HeLa细胞,A549细胞,SW480细胞和HL-7702细胞)的端粒中的不同氧化损伤水平图;其中,基因组DNA的量为50ng。误差棒为三组实验的标准偏差。Figure 10 is a graph showing different oxidative damage levels in telomeres from different human cell lines (i.e., HeLa cells, A549 cells, SW480 cells, and HL-7702 cells) treated with 1000 μMH 2 O2 , wherein the amount of genomic DNA is 50 ng. Error bars are standard deviations of three sets of experiments.
具体实施方式DETAILED DESCRIPTION
应该指出,以下详细说明都是例示性的,旨在对本申请提供进一步的说明。除非另有指明,本文使用的所有技术和科学术语具有与本申请所属技术领域的普通技术人员通常理解的相同含义。It should be noted that the following detailed descriptions are illustrative and are intended to provide further explanation of the present application. Unless otherwise specified, all technical and scientific terms used herein have the same meanings as those commonly understood by those skilled in the art to which the present application belongs.
需要注意的是,这里所使用的术语仅是为了描述具体实施方式,而非意图限制根据本申请的示例性实施方式。如在这里所使用的,除非上下文另外明确指出,否则单数形式也意图包括复数形式,此外,还应当理解的是,当在本说明书中使用术语“包含”和/或“包括”时,其指明存在特征、步骤、操作、器件、组件和/或它们的组合。It should be noted that the terms used herein are only for describing specific embodiments and are not intended to limit the exemplary embodiments according to the present application. As used herein, unless the context clearly indicates otherwise, the singular form is also intended to include the plural form. In addition, it should be understood that when the terms "comprising" and/or "including" are used in this specification, it indicates the presence of features, steps, operations, devices, components and/or combinations thereof.
如前所述,目前检测基因组中8-oxoG的最常用方法包括高效液相色谱(HPLC)与电化学检测相结合(HPLC-ECD)以及HPLC与气相色谱-质谱联用(HPLC-MS)/MS)。这些方法可以提供更准确的8-oxoG计数,但需要把DNA序列消化成单个核苷酸碱基,而且需要复杂的仪器设备及繁琐的实验步骤。迄今为止,由于低丰度和高度重复的基因区域,很少有方法可用于检测端粒中的氧化性损伤水平。As mentioned above, the most commonly used methods for detecting 8-oxoG in the genome include high performance liquid chromatography (HPLC) coupled with electrochemical detection (HPLC-ECD) and HPLC coupled with gas chromatography-mass spectrometry (HPLC-MS/MS). These methods can provide more accurate 8-oxoG counts, but require digestion of DNA sequences into single nucleotide bases and require complex instrumentation and cumbersome experimental steps. To date, few methods have been available for detecting oxidative damage levels in telomeres due to the low abundance and highly repetitive gene regions.
有鉴于此,本发明的一个具体实施方式中,提供一种检测端粒内氧化性损伤的荧光化学传感器,所述荧光化学传感器至少包括信号探针、捕获探针和辅助探针。In view of this, in a specific embodiment of the present invention, a fluorescent chemical sensor for detecting oxidative damage in telomeres is provided, wherein the fluorescent chemical sensor at least includes a signal probe, a capture probe and an auxiliary probe.
本发明的又一具体实施方式中,所述信号探针为具有脱嘌呤嘧啶位点(AP位点)的富T单链DNA序列,所述信号探针5'端修饰有生物素,所述信号探针3'端修饰有荧光基团,优选使用AF488进行修饰;In another specific embodiment of the present invention, the signal probe is a T-rich single-stranded DNA sequence with an apurinic pyrimidine site (AP site), the 5' end of the signal probe is modified with biotin, and the 3' end of the signal probe is modified with a fluorescent group, preferably AF488;
本发明的又一具体实施方式中,所述信号探针与链霉抗生物素蛋白包被的磁珠进行缀合。In another specific embodiment of the present invention, the signal probe is conjugated to magnetic beads coated with streptavidin.
本发明的又一具体实施方式中,所述信号探针的碱基序列如下:In another specific embodiment of the present invention, the base sequence of the signal probe is as follows:
5'-Biotin-TTT TTT TTT XTT TTT TTT-AF488-3';5'-Biotin-TTT TTT TTT XTT TTT TTT-AF488-3';
其中X代表脱嘌呤嘧啶位点;Where X represents an apurinic pyrimidine site;
所述捕获探针用于捕获富集端粒,其具有与端粒杂交的单链DNA序列,所述捕获探针3'端进行生物素化修饰;The capture probe is used to capture and enrich telomeres, and has a single-stranded DNA sequence that hybridizes with the telomeres. The 3' end of the capture probe is biotinylated;
本发明的又一具体实施方式中,所述捕获探针的碱基序列如下:In another specific embodiment of the present invention, the base sequence of the capture probe is as follows:
CCT AAC CCT AAC CCT AAC CCT TTT-Biotin;CCT AAC CCT AAC CCT AAC CCT TTT-Biotin;
本发明的又一具体实施方式中,所述辅助探针为3'端用ddC修饰的单链DNA序列,其与单链端粒杂交形成甲酰胺嘧啶[fapy]-DNA糖基化酶的dsDNA底物。In another specific embodiment of the present invention, the auxiliary probe is a single-stranded DNA sequence modified with ddC at the 3' end, which hybridizes with the single-stranded telomere to form a dsDNA substrate for formamidopyrimidine [fapy]-DNA glycosylase.
本发明的又一具体实施方式中,所述辅助探针的碱基序列如下:In another specific embodiment of the present invention, the base sequence of the auxiliary probe is as follows:
5'-CCT AAC CCT AAC CCT AAddC-3'。5'-CCT AAC CCT AAC CCT AAddC-3'.
本发明的又一具体实施方式中,所述荧光化学传感器还包括甲酰胺嘧啶[fapy]-DNA糖基化酶、末端脱氧核苷酸转移酶(TdT)、核酸内切酶IV和T4-多核苷酸激酶。In another specific embodiment of the present invention, the fluorescent chemical sensor further comprises formamidopyrimidine [fapy]-DNA glycosylase, terminal deoxynucleotidyl transferase (TdT), endonuclease IV and T4-polynucleotide kinase.
本发明的又一具体实施方式中,所述端粒内氧化性损伤包括8-oxoG损伤。In another specific embodiment of the present invention, the oxidative damage in the telomere includes 8-oxoG damage.
本发明的又一具体实施方式中,提供基于上述荧光化学传感器检测端粒内氧化性损伤的方法,所述方法包括:In another specific embodiment of the present invention, a method for detecting oxidative damage in telomeres based on the above-mentioned fluorescent chemical sensor is provided, and the method comprises:
S1、用dsDNA片段酶消化提取基因组DNA,获得具有50-200bp的DNA;S1. Extract genomic DNA by digestion with dsDNA fragmentase to obtain DNA with 50-200 bp;
S2、加入捕获探针孵育处理,用于捕获富集端粒;S2, adding capture probes for incubation to capture and enrich telomeres;
S3、加入TdT进行孵育处理,催化三磷酸脱氧核苷酸(dATPs)重复加入DNA分子的3'-羟基末端(3'-OH);S3, adding TdT for incubation treatment, catalyzing the repeated addition of deoxynucleotide triphosphates (dATPs) to the 3'-hydroxyl end (3'-OH) of the DNA molecule;
S4、加入辅助探针、甲酰胺嘧啶[fapy]-DNA糖基化酶进行孵育处理,其中辅助探针用于与单链端粒杂交以形成甲酰胺嘧啶[fapy]-DNA糖基化酶的dsDNA底物;S4, adding an auxiliary probe and fapy-DNA glycosylase for incubation, wherein the auxiliary probe is used to hybridize with the single-stranded telomere to form a dsDNA substrate for fapy-DNA glycosylase;
S5、加入TdT、核酸内切酶IV和磁珠偶联信号探针进行孵育处理;S5, adding TdT, endonuclease IV and magnetic bead-coupled signal probe for incubation;
S6、磁分离后,对上清液进行单分子检测。S6. After magnetic separation, the supernatant is subjected to single molecule detection.
本发明的又一具体实施方式中,所述步骤S2中,孵育处理条件为于90~100℃(优选95℃)温育3~8分钟(优选5分钟);In another specific embodiment of the present invention, in step S2, the incubation treatment condition is incubating at 90-100° C. (preferably 95° C.) for 3-8 minutes (preferably 5 minutes);
本发明的又一具体实施方式中,所述步骤S3中,孵育处理过程中还加入ddCTP,防止端粒发生非特异性扩增;In another specific embodiment of the present invention, in step S3, ddCTP is also added during the incubation process to prevent non-specific amplification of telomeres;
孵育处理条件为于30~40℃(优选37℃)孵育20~40分钟(优选30分钟);The incubation treatment conditions are 30-40°C (preferably 37°C) for 20-40 minutes (preferably 30 minutes);
本发明的又一具体实施方式中,所述步骤S4中,孵育处理条件为30~40℃(优选为37℃)下孵育40~80分钟(优选为60分钟);In another specific embodiment of the present invention, in step S4, the incubation treatment condition is 30-40° C. (preferably 37° C.) for 40-80 minutes (preferably 60 minutes);
本发明的又一具体实施方式中,所述步骤S5中,孵育处理条件为30~40℃(优选为37℃)下孵育40~80分钟(优选为60分钟);In another specific embodiment of the present invention, in step S5, the incubation treatment condition is 30-40° C. (preferably 37° C.) for 40-80 minutes (preferably 60 minutes);
本发明的又一具体实施方式中,所述步骤S6中,采用全内反射荧光(TIRF)技术进行单分子检测;同时,通过荧光分光光度计测量反应产物的荧光光谱;具体为:在488nm的激发波长下测量AF488荧光,并使用532nm的荧光强度进行定量分析。In another specific embodiment of the present invention, in step S6, total internal reflection fluorescence (TIRF) technology is used for single molecule detection; at the same time, the fluorescence spectrum of the reaction product is measured by a fluorescence spectrophotometer; specifically: AF488 fluorescence is measured at an excitation wavelength of 488nm, and the fluorescence intensity at 532nm is used for quantitative analysis.
本发明的又一具体实施方式中,提供上述荧光化学传感器和/或检测方法在检测端粒内氧化性损伤中的应用。所述端粒内氧化性损伤包括8-oxoG损伤。In another specific embodiment of the present invention, the use of the above fluorescent chemical sensor and/or detection method in detecting oxidative damage in telomeres is provided. The oxidative damage in telomeres includes 8-oxoG damage.
以下通过实施例对本发明做进一步解释说明,但不构成对本发明的限制。应理解这些实施例仅用于说明本发明而不用于限制本发明的范围。另外,实施例中未详细说明的分子生物学方法均为本领域常规的方法,具体操作可参看分子生物指南或产品说明书。The present invention is further explained by the following examples, but it does not constitute a limitation of the present invention. It should be understood that these examples are only used to illustrate the present invention and are not used to limit the scope of the present invention. In addition, the molecular biology methods not described in detail in the examples are all conventional methods in the art, and the specific operations can be referred to the molecular biology guide or product instructions.
实施例1Example 1
1.本发明检测原理及方法步骤:1. Detection principle and method steps of the present invention:
首先用dsDNA片段酶消化提取基因组DNA,以获得具有50-200bp的DNA。在人类基因组中,端粒由重复序列(TTAGGG)n组成,其形成终止于单链3'G-富集突出端的双链DNA的长区域。为了富集端粒,3'-生物素化捕获探针设计用于与端粒杂交并利用链霉抗生物素蛋白包被的磁珠将端粒从基因组DNA中分离出来。所有分离的端粒通过使用TdT聚合用ddCTP进行末端标记,以防止非特异性扩增。随后,为了提高碱基切除修复反应的效率,还设计了在3'末端用ddC修饰的辅助探针与单链端粒杂交以形成甲酰胺嘧啶[fapy]-DNA糖基化酶(Fpg)的dsDNA底物。First, genomic DNA was extracted by digestion with dsDNA fragmentase to obtain DNA with 50-200bp. In the human genome, telomeres are composed of repetitive sequences (TTAGGG)n , which form long regions of double-stranded DNA terminating in single-stranded 3'G-enriched overhangs. In order to enrich telomeres, 3'-biotinylated capture probes are designed to hybridize with telomeres and separate telomeres from genomic DNA using streptavidin-coated magnetic beads. All separated telomeres are end-labeled with ddCTP by using TdT polymerization to prevent nonspecific amplification. Subsequently, in order to improve the efficiency of the base excision repair reaction, an auxiliary probe modified with ddC at the 3' end was also designed to hybridize with single-stranded telomeres to form a dsDNA substrate for formamidopyrimidine [fapy]-DNA glycosylase (Fpg).
该方法涉及三个连续步骤:(1)在辅助探针的辅助下,Fpg诱导8-oxoG碱基切除修复反应,(2)TdT介导的延伸反应激活的Endo IV-辅助信号探针的循环切割,和(3)释放的AF488荧光团的单分子检测。The method involves three sequential steps: (1) Fpg-induced 8-oxoG base excision repair reaction with the assistance of an auxiliary probe, (2) cyclic cleavage of the Endo IV-auxiliary signal probe activated by a TdT-mediated extension reaction, and (3) single-molecule detection of the released AF488 fluorophore.
来自大肠杆菌的Fpg含有DNA糖基化酶活性和β,δ-消除AP裂解酶活性,可以去除dsDNA中受损的氧化性损伤碱基并切割DNA骨架,与5'-磷酸和3'-磷酸酯残留物形成单核苷酸缺口。如图1所示,端粒-辅助探针杂交体中受损的氧化性损伤碱基可以通过Fpg除去,在5'和3'磷酸基团的修饰位点产生核苷酸缺口,导致端粒和辅助探针的解离。切割的端粒的3'-磷酸基团可以通过多核苷酸激酶(PNK)除去以产生游离的3'-OH末端,其随后用作引物以启动TdT催化的模板独立的聚合延伸以产生在dATP存在下的聚腺苷(polyA)。设计了磁珠(MB)-缀合的富含T的信号探针,其在序列的中间用AP位点修饰,并在3'末端用AF488标记。得到的poly-A序列可以与MB连接的富含T的信号探针的杂交以形成具有完整AP位点的polyA-信号探针DNA双链体,其可以被Endo IV切割,导致信号探针的断裂和释放polyA序列,AF488荧光团和切割的含有游离的3'-OH末端的MB共轭信号探针。释放的polyA序列可与新信号探针结合形成具有完整AP位点的新dsDNA,从而启动Endo IV诱导的polyA-信号探针双链体的环状切割和更多AF488荧光团的释放。此外,具有游离3'-OH末端的裂解的MB-缀合的信号探针可以作为引物在dATP存在下引发新的TdT介导的延伸并产生大量的长polyA产物。得到的MB连接的polyA产物可以与多个MB缀合的信号探针杂交以形成具有完整AP位点的新dsDNA,其启动Endo IV诱导的polyA产物-信号探针双链体的循环切割,释放出更多AF488荧光团,MB-连接的polyA产物和具有游离的3'-OH末端的MB-缀合的信号探针。因此,所有释放的polyA产物和具有游离3'-OH末端的MB缀合的信号探针可以诱导信号探针的多次循环切割和更多AF488荧光团的释放,直到酶没有活性。最后,释放的AF488荧光团可以通过磁分离后的单分子检测进行简单定量。Fpg from Escherichia coli contains DNA glycosylase activity and β,δ-eliminating AP lyase activity, which can remove damaged oxidatively damaged bases in dsDNA and cut the DNA backbone to form single nucleotide gaps with 5'-phosphate and 3'-phosphate residues. As shown in Figure 1, damaged oxidatively damaged bases in the telomere-auxiliary probe hybrid can be removed by Fpg, generating nucleotide gaps at the modification sites of the 5' and 3' phosphate groups, leading to the dissociation of the telomere and the auxiliary probe. The 3'-phosphate group of the cut telomere can be removed by polynucleotide kinase (PNK) to generate a free 3'-OH end, which is then used as a primer to initiate TdT-catalyzed template-independent polymerization extension to produce polyadenosine (polyA) in the presence of dATP. A magnetic bead (MB)-conjugated T-rich signal probe was designed, which was modified with an AP site in the middle of the sequence and labeled with AF488 at the 3' end. The resulting poly-A sequence can hybridize with the MB-linked T-rich signal probe to form a polyA-signal probe DNA duplex with an intact AP site, which can be cleaved by Endo IV, resulting in the breakage of the signal probe and the release of the polyA sequence, the AF488 fluorophore, and the cleaved MB-conjugated signal probe containing a free 3'-OH terminus. The released polyA sequence can bind to the new signal probe to form a new dsDNA with an intact AP site, thereby initiating Endo IV-induced circular cleavage of the polyA-signal probe duplex and the release of more AF488 fluorophores. In addition, the cleaved MB-conjugated signal probe with a free 3'-OH terminus can serve as a primer to initiate a new TdT-mediated extension in the presence of dATP and generate a large amount of long polyA products. The resulting MB-linked polyA product can hybridize with multiple MB-conjugated signal probes to form new dsDNA with intact AP sites, which initiates Endo IV-induced cyclic cleavage of the polyA product-signal probe duplex, releasing more AF488 fluorophores, MB-linked polyA products, and MB-conjugated signal probes with free 3'-OH termini. Therefore, all released polyA products and MB-conjugated signal probes with free 3'-OH termini can induce multiple cyclic cleavages of the signal probes and the release of more AF488 fluorophores until the enzyme is inactive. Finally, the released AF488 fluorophores can be simply quantified by single-molecule detection after magnetic separation.
信号探针与链霉抗生物素蛋白包被的磁珠的缀合:Conjugation of signal probe to streptavidin-coated magnetic beads:
根据Invitrogen Corporation的方案进行信号探针与磁珠(MB)的偶联。对于信号探针,将100μL链霉抗生物素蛋白包被的MBs溶液(10mg/mL)转移到600μL离心管中,用1×B&W缓冲液(5mM Tris-HCl(pH 7.5),0.5mM EDTA,1M NaCl)洗涤两次。通过磁力分离除去上清液,并将MB重悬于2×B&W缓冲液(10mM Tris-HCl(pH 7.5),1mM EDTA,2M NaCl)中至终浓度为5μg/μL。然后将200μL的1μM生物素化的信号探针与200μL的5μg/μLMBs溶液混合,在室温下在辊式混合器中在黑暗中温育10分钟。然后将混合物洗涤三次以除去未偶联的信号探针,并将剩余的信号探针-MB缀合物重悬于100μL的TE缓冲液中。The coupling of the signal probe to magnetic beads (MB) was performed according to the protocol of Invitrogen Corporation. For the signal probe, 100 μL of the streptavidin-coated MBs solution (10 mg/mL) was transferred to a 600 μL centrifuge tube and washed twice with 1×B&W buffer (5 mM Tris-HCl (pH 7.5), 0.5 mM EDTA, 1 M NaCl). The supernatant was removed by magnetic separation, and the MB was resuspended in 2×B&W buffer (10 mM Tris-HCl (pH 7.5), 1 mM EDTA, 2 M NaCl) to a final concentration of 5 μg/μL. Then 200 μL of 1 μM biotinylated signal probe was mixed with 200 μL of 5 μg/μL MBs solution and incubated in the dark on a roller mixer at room temperature for 10 minutes. The mixture was then washed three times to remove the uncoupled signal probe, and the remaining signal probe-MB conjugate was resuspended in 100 μL of TE buffer.
8-oxoG碱基切除修复和TdT介导的延伸反应激活的信号探针的Endo IV-辅助切割。为了切除8-oxoG,将含有各种浓度的含有8-oxoG损伤的合成端粒,10pM辅助探针,1x末端转移酶反应缓冲液,1.6U Fpg和3U T4-多核苷酸激酶(PNK)的反应混合物(10μL)在37℃中温育1小时。然后将4U TdT,5U核酸内切酶IV,1x末端转移酶反应缓冲液,0.25mMCoCl2,250μM dATP和磁珠偶联信号探针加入到反应混合物中,总体积为50μL,然后在37℃下孵育60分钟。最后,通过磁力分离将链霉抗生物素蛋白包被的MB分离3分钟,并对上清液进行单分子检测。Endo IV-assisted cleavage of the signal probe activated by 8-oxoG base excision repair and TdT-mediated extension reaction. To excise 8-oxoG, a reaction mixture (10 μL) containing various concentrations of synthetic telomeres containing 8-oxoG lesions, 10 pM auxiliary probe, 1× terminal transferase reaction buffer, 1.6 U Fpg, and 3 U T4-polynucleotide kinase (PNK) was incubated at 37°C for 1 hour. Then 4 U TdT, 5 U endonuclease IV, 1× terminal transferase reaction buffer, 0.25 mM CoCl2 , 250 μM dATP, and magnetic bead-coupled signal probe were added to the reaction mixture in a total volume of 50 μL, followed by incubation at 37°C for 60 minutes. Finally, streptavidin-coated MBs were separated by magnetic separation for 3 minutes, and the supernatant was subjected to single-molecule detection.
基于全内反射荧光(TIRF)的单分子检测:Single molecule detection based on total internal reflection fluorescence (TIRF):
通过全内反射荧光显微镜(TIRF)(Nikon,Ti-E,Japan)获得单分子的图像。稀释1000倍后,将10μL反应产物样品用于TIRF成像,488nm激光器用于激发AF488。使用油浸100×物镜收集光子。来自AF488的光子通过相机收集(Photometrics,Evolve 512)。对于数据分析,使用Image J软件选择600×600像素的成像区域用于AF488分子计数。通过计算十帧获得平均AF488计数。Single molecule images were obtained by total internal reflection fluorescence microscopy (TIRF) (Nikon, Ti-E, Japan). After
荧光光谱的测量:Fluorescence spectrum measurement:
通过Hitachi F-7000荧光分光光度计(Tokyo,Japan)测量50μL反应产物的荧光光谱。在488nm的激发波长下测量AF488荧光,并使用532nm的荧光强度进行定量分析。The fluorescence spectrum of 50 μL of the reaction product was measured by a Hitachi F-7000 fluorescence spectrophotometer (Tokyo, Japan). AF488 fluorescence was measured at an excitation wavelength of 488 nm, and the fluorescence intensity at 532 nm was used for quantitative analysis.
凝胶电泳分析:Gel electrophoresis analysis:
通过14%非变性聚丙烯酰胺凝胶电泳在1×Tris-硼酸盐-EDTA(TBE)缓冲液(9mMTris-HCl,9mM硼酸,0.2mM EDTA,pH7.9)中在110V下分析用SYBR Gold染色的产物。在室温下恒定电压40分钟。通过ChemiDoc MP成像系统(Hercules,CA,U.S.A。)使凝胶电泳图像可视化。Products stained with SYBR Gold were analyzed by 14% non-denaturing polyacrylamide gel electrophoresis in 1× Tris-borate-EDTA (TBE) buffer (9 mM Tris-HCl, 9 mM boric acid, 0.2 mM EDTA, pH 7.9) at 110 V. Constant voltage was maintained at room temperature for 40 minutes. Gel electrophoresis images were visualized by ChemiDoc MP imaging system (Hercules, CA, U.S.A.).
细胞培养和基因组DNA的制备:Cell culture and genomic DNA preparation:
将细胞在含有10%胎牛血清(FBS,Invitrogen,USA)的Dulbecco改良的Eagle培养基(DMEM;Gibco,U.S.A。)中于37℃,5%CO2下培养。为了在体内诱导氧化性DNA损伤,用PBS缓冲液洗涤细胞,并在37℃,5%CO2的潮湿室中用不同浓度的H2O2处理1小时。按照制造商的建议,使用
检测HeLa细胞端粒内8-oxoG损伤:Detection of 8-oxoG damage in HeLa cell telomeres:
为了从基因组DNA中富集端粒,将生物素化的捕获探针和消化的基因组DNA在1×末端转移酶反应缓冲液中于95℃温育5分钟,然后缓慢冷却至室温。然后通过磁珠(MB)分离生物素化的捕获探针。磁力分离并洗涤三次后,加入4U TdT,200μM ddCTP,1×末端转移酶反应缓冲液和0.25mM CoCl2,总体积为10μL,并在37℃下孵育30分钟。通过磁力分离除去过量的TdT和ddCTP。然后将100nM辅助探针,1x末端转移酶反应缓冲液,3U甲酰胺嘧啶[fapy]-DNA糖基化酶(Fpg),5U T4-多核苷酸激酶加入到反应体系中,总体积为10μL,并在37℃下孵育1小时。然后将4U TdT,5U核酸内切酶IV,1×末端转移酶反应缓冲液,0.25mM CoCl2,250μM dATP和磁珠偶联信号探针加入到反应混合物中,总体积为50μL,然后在37℃下孵育60分钟。磁分离后,按照上述步骤进行单分子检测。To enrich telomeres from genomic DNA, the biotinylated capture probe and digested genomic DNA were incubated in 1× terminal transferase reaction buffer at 95°C for 5 minutes and then slowly cooled to room temperature. The biotinylated capture probe was then separated by magnetic beads (MB). After magnetic separation and three washes, 4U TdT, 200μM ddCTP, 1× terminal transferase reaction buffer, and 0.25mM CoCl2 were added in a total volume of 10μL and incubated at 37°C for 30 minutes. Excess TdT and ddCTP were removed by magnetic separation. Then 100nM auxiliary probe, 1x terminal transferase reaction buffer, 3U formamidopyrimidine [fapy]-DNA glycosylase (Fpg), 5U T4-polynucleotide kinase were added to the reaction system in a total volume of 10μL and incubated at 37°C for 1 hour. Then 4U TdT, 5U endonuclease IV, 1× terminal transferase reaction buffer, 0.25mM CoCl2 , 250μM dATP and magnetic bead-coupled signal probe were added to the reaction mixture in a total volume of 50μL, and then incubated at 37°C for 60 minutes. After magnetic separation, single molecule detection was performed according to the above steps.
实验所需核苷酸序列Nucleotide sequences required for the experiment


试验结果分析Test results analysis
1.检测可行性实验1. Feasibility test
氧化性碱基损伤,尤其是8-oxoG和FapyG是最常见的氧化性DNA损伤。使用含有一个8-oxoG碱基(Telo-Cy5)的Cy5标记的合成端粒来验证Fpg可以消除氧化性DNA损伤和切割DNA骨架以产生新的切割产物(图6)。然后选择含有一个8-oxoG(Telo-1)的合成端粒来验证氧化损伤的去除是否可以诱导TdT介导的聚合反应。进行了非变性聚丙烯酰胺凝胶电泳(PAGE)来分析扩增产物(图2A)。在PNK+TdT存在的情况下,仅观察到Telo-1/辅助探针双链的条带(图2A,泳道1)。当存在Fpg,PNK和TdT酶时,观察到明确定义的扩增产物条带(图2A,泳道2),表明Fpg可从端粒中除去8-oxoG碱基并启动随后的TdT催化的聚合延伸。此外,使用含有两个8-oxoG碱基(Telo-2)的合成端粒作为目标来研究所提出的氧化损伤检测方法的可行性(图2B和C)。如图2B所示,在Telo-2存在下观察到明显的AF488荧光信号(图2B,红线)。相反,在没有氧化损伤(Telo-0)的正常端粒中没有检测到明显的AF488信号(图2B,黑线)。通过单分子计数进一步证实了这些结果(图2C)。观察到Telo-2对应的不同的AF488荧光信号(图2C,I),但没有检测到响应于Telo-0的AF488荧光信号(图2C,II)。Oxidative base damage, especially 8-oxoG and FapyG, is the most common oxidative DNA damage. A Cy5-labeled synthetic telomere containing an 8-oxoG base (Telo-Cy5) was used to verify that Fpg can eliminate oxidative DNA damage and cleave the DNA backbone to generate new cleavage products (Figure 6). Then a synthetic telomere containing an 8-oxoG (Telo-1) was selected to verify whether the removal of oxidative damage can induce TdT-mediated polymerization. Non-denaturing polyacrylamide gel electrophoresis (PAGE) was performed to analyze the amplification products (Figure 2A). In the presence of PNK+TdT, only the band of the Telo-1/auxiliary probe double strand was observed (Figure 2A, lane 1). When Fpg, PNK and TdT enzymes were present, well-defined amplification product bands were observed (Figure 2A, lane 2), indicating that Fpg can remove the 8-oxoG base from the telomere and initiate subsequent TdT-catalyzed polymerization extension. In addition, synthetic telomeres containing two 8-oxoG bases (Telo-2) were used as targets to investigate the feasibility of the proposed oxidative damage detection method (Figures 2B and C). As shown in Figure 2B, a clear AF488 fluorescence signal was observed in the presence of Telo-2 (Figure 2B, red line). In contrast, no obvious AF488 signal was detected in normal telomeres without oxidative damage (Telo-0) (Figure 2B, black line). These results were further confirmed by single molecule counting (Figure 2C). Different AF488 fluorescence signals corresponding to Telo-2 were observed (Figure 2C, I), but no AF488 fluorescence signal was detected in response to Telo-0 (Figure 2C, II).
2.选择性的检测2. Selective detection
氧化性DNA损伤的位置在整个基因组中可能是随机的。为了研究所提出的方法检测端粒氧化损伤的选择性,使用含有两个8-oxoG碱基的端粒序列(Telo-2)和含有两个8-oxoG碱基的合成KRAS序列(KRAS),随机DNA(Random DNA)作为对照。将端粒特异性捕获探针与磁珠组合分别用于分离Telo-2,KRAS和Random DNA,然后通过所提出的方法检测氧化损伤。在相同条件下,Telo-2观察到高荧光信号,而对于KRAS,RandomDNA和仅具有反应缓冲液的对照组没有观察到明显的荧光信号(图3)。这可以通过端粒特异性捕获探针仅捕获端粒的事实来解释。这些结果清楚地表明,所提出的方法可用于选择性检测端粒中的氧化损伤。The location of oxidative DNA damage may be random throughout the genome. In order to investigate the selectivity of the proposed method for detecting telomere oxidative damage, a telomere sequence containing two 8-oxoG bases (Telo-2) and a synthetic KRAS sequence containing two 8-oxoG bases (KRAS), random DNA (Random DNA) were used as controls. Telomere-specific capture probes in combination with magnetic beads were used to separate Telo-2, KRAS and Random DNA, respectively, and then oxidative damage was detected by the proposed method. Under the same conditions, a high fluorescence signal was observed for Telo-2, while no obvious fluorescence signal was observed for KRAS, RandomDNA and the control group with only reaction buffer (Figure 3). This can be explained by the fact that the telomere-specific capture probe only captures telomeres. These results clearly show that the proposed method can be used to selectively detect oxidative damage in telomeres.
3.单分子检测低丰度端粒内8-oxoG损伤3. Single-molecule detection of low-abundance 8-oxoG lesions in telomeres
在最佳实验条件下(图7和图4),使用合成的Telo-2来研究所提出的氧化损伤检测方法的灵敏度。如图4A所示,AF488计数与氧化损伤浓度在1×10-16至1×10-11M的5个数量级范围内呈现对数线性相关。回归方程为N=1632.6+101.16log10C,相关系数为0.984,其中N是测量的AF488计数,C是氧化损伤浓度(Telo-2)。通过评估阴性对照的平均响应值加上标准偏差的三倍,计算出检测限为9.3×10-17M。与基于毛细管电泳的荧光法(2.9×10-12M)相比,该方法的灵敏度提高了4个数量级,与基于适体的传感器的(3×10-9M)方法相比提高了7个数量级。此外,检测了不同比例的含有两个8-oxoG碱基的Telo-2和正常端粒(Telo-0)的混合物中的氧化损伤水平。AF488计数随着混合物中输入氧化损伤量的增加而增强(图9),并且在测量的氧化损伤水平与实际输入氧化损伤水平之间获得线性关系(图4B)。该结果表明,所提出的方法可以量化低至0.001%的氧化损伤水平,而不受复杂样品中大量正常DNA序列的干扰,优于报道的基于qPCR的测定。该方法的高灵敏度可归因于以下因素:(1)Fpg可有效去除端粒中的氧化损伤;(2)只有由切除的氧化损伤位点产生的游离3'-OH才能引发TdT诱导的聚合反应,导致极低的背景信号;(3)TdT介导的聚合延伸可以诱导Endo IV辅助的信号探针的循环切割,显著放大荧光信号;(4)引入磁分离和单分子检测,进一步降低了背景信号,提高了检测灵敏度。Under the optimal experimental conditions (Figures 7 and 4), the sensitivity of the proposed oxidative damage detection method was investigated using synthetic Telo-2. As shown in Figure 4A, the AF488 counts showed a log-linear correlation with the oxidative damage concentration over a five-order-of-magnitude range from 1×10-16 to 1×10-11 M. The regression equation was N=1632.6+101.16log10 C, with a correlation coefficient of 0.984, where N was the measured AF488 counts and C was the oxidative damage concentration (Telo-2). The detection limit was calculated to be 9.3×10-17 M by evaluating the mean response value of the negative control plus three times the standard deviation. The sensitivity of this method was improved by 4 orders of magnitude compared to the capillary electrophoresis-based fluorescence method (2.9×10-12 M) and by 7 orders of magnitude compared to the aptamer-based sensor (3×10-9 M). In addition, the oxidative damage levels in mixtures of Telo-2 containing two 8-oxoG bases and normal telomeres (Telo-0) at different ratios were detected. The AF488 counts increased with the increase in the amount of input oxidative damage in the mixture (Figure 9), and a linear relationship was obtained between the measured oxidative damage level and the actual input oxidative damage level (Figure 4B). This result indicates that the proposed method can quantify oxidative damage levels as low as 0.001% without interference from a large amount of normal DNA sequences in complex samples, which is superior to the reported qPCR-based assays. The high sensitivity of this method can be attributed to the following factors: (1) Fpg can effectively remove oxidative damage in telomeres; (2) only free 3'-OH generated by the excised oxidative damage site can trigger TdT-induced polymerization, resulting in extremely low background signals; (3) TdT-mediated polymerization extension can induce cyclic cleavage of Endo IV-assisted signal probes, significantly amplifying the fluorescence signal; (4) The introduction of magnetic separation and single-molecule detection further reduced the background signal and improved the detection sensitivity.
4.检测由活性氧引起的基因组DNA端粒内的氧化性损伤4. Detection of oxidative damage in genomic DNA telomeres caused by reactive oxygen species
该方法测量了从暴露于不同浓度的过氧化氢(H2O2)的人宫颈癌细胞系(HeLa细胞)中提取的端粒氧化损伤水平。人端粒含有数千个碱基对和100~200个核苷酸单个3'末端的链状突出端,可折叠成独特的二级结构(即Gquadruplex)。基因组DNA中分离的端粒含量估计为0.018%-0.032%(表1)。随着H2O2浓度从0增加到1000μM,AF488计数增强,表明氧化损伤与H2O2浓度的增加成正比(图5A)。该方法进一步测量了来自用1000μM H2O2处理的HeLa细胞的不同量的基因组DNA中分离的端粒的氧化损伤。如图5B所示,AF488计数随着基因组DNA量的增加而提高,并且在AF488计数和基因组DNA量的对数之间获得0.1至100ng范围内的线性相关性。相应的方程是N=94.02+82.28log10A,相关系数为0.97,其中N代表AF488计数,A代表基因组DNA的数量。通过评估阴性对照的平均响应加上标准偏差的三倍,计算检测限为0.078ng。重要的是,根据标准曲线(N=1632.6+101.16log10C,图4A),计算出每个细胞端粒含有为34-44个氧化性损伤碱基(表2)。值得注意的是,与基于HPLC的方法(25-50μg)相比,该方法具有仅涉及少量DNA(0.1ng)的显着优点。并且进一步测量了不同细胞系中端粒的氧化损伤水平。癌细胞中的氧化损伤水平高于正常细胞(图10)。The method measured the level of oxidative damage in telomeres extracted from a human cervical cancer cell line (HeLa cells) exposed to different concentrations of hydrogen peroxide (H2 O2 ). Human telomeres contain thousands of base pairs and a chain-like overhang at the single 3' end of 100-200 nucleotides, which can be folded into a unique secondary structure (i.e., Gquadruplex). The content of telomeres isolated from genomic DNA was estimated to be 0.018%-0.032% (Table 1). As the concentration of H2 O2 increased from 0 to 1000 μM, the AF488 counts increased, indicating that the oxidative damage was proportional to the increase in the concentration of H2 O2 (Figure 5A). The method further measured the oxidative damage of telomeres isolated from different amounts of genomic DNA from HeLa cells treated with 1000 μM H2 O2. As shown in Figure 5B, the AF488 counts increased with the increase in the amount of genomic DNA, and a linear correlation was obtained between the AF488 counts and the logarithm of the amount of genomic DNA in the range of 0.1 to 100 ng. The corresponding equation is N=94.02+82.28log10 A, with a correlation coefficient of 0.97, where N represents AF488 counts and A represents the amount of genomic DNA. The detection limit was calculated to be 0.078 ng by evaluating the mean response of the negative control plus three times the standard deviation. Importantly, based on the standard curve (N=1632.6+101.16log10 C, Figure 4A), it was calculated that each cell telomere contained 34-44 oxidatively damaged bases (Table 2). It is worth noting that this method has the significant advantage of involving only a small amount of DNA (0.1 ng) compared to the HPLC-based method (25-50 μg). And the level of oxidative damage to telomeres in different cell lines was further measured. The level of oxidative damage in cancer cells is higher than that in normal cells (Figure 10).
表1.分离的端粒含量来自基因组DNATable 1. Telomere content isolated from genomic DNA
表2.计算的每个细胞中端粒的氧化损伤数Table 2. Calculated number of telomere oxidative damage per cell

5.验证Fpg介导的氧化损伤的去除5. Validation of Fpg-mediated Removal of Oxidative Damage
使用含有一个8-oxoG碱基(Telo-Cy5)的Cy5标记的合成端粒来验证Fpg是否可以去除氧化性DNA损伤。辅助探针可以与Telo-Cy5杂交形成Telo-Cy5/辅助探针双链体。非变性聚丙烯酰胺凝胶电泳(PAGE)分析显示在没有Fpg的情况下仅观察到Telo-Cy5/辅助探针双链体的条带(图6,泳道1),表明没有8-oxoG被切割。在存在Fpg的情况下,观察到对应于裂解产物大小的新条带(图6,泳道2),表明Fpg可以去除Telo-Cy5中的8-oxoG碱基损伤并切割DNA骨架以产生一种新的裂解产物。A Cy5-labeled synthetic telomere containing an 8-oxoG base (Telo-Cy5) was used to verify whether Fpg can remove oxidative DNA damage. The auxiliary probe can hybridize with Telo-Cy5 to form a Telo-Cy5/auxiliary probe duplex. Non-denaturing polyacrylamide gel electrophoresis (PAGE) analysis showed that only bands of the Telo-Cy5/auxiliary probe duplex were observed in the absence of Fpg (Figure 6, lane 1), indicating that no 8-oxoG was cut. In the presence of Fpg, a new band corresponding to the size of the cleavage product was observed (Figure 6, lane 2), indicating that Fpg can remove the 8-oxoG base damage in Telo-Cy5 and cut the DNA backbone to produce a new cleavage product.
6.实验条件的优化6. Optimization of Experimental Conditions
为了获得最佳的分析性能,分别优化了Endo IV和TdT的量。如图7所示,当TdT的量固定在8U时,AF488计数随着的增加而增加,并且Endo IV的量到4U时达到最大值。因此,4U的Endo IV用于后续研究。如图8所示,当Endo IV的量固定在4U时,AF488计数随着的增加而增加,并且TdT量到6U时达到最大值。因此,使用6U的TdT在随后的研究中。In order to obtain the best analytical performance, the amounts of Endo IV and TdT were optimized, respectively. As shown in Figure 7, when the amount of TdT was fixed at 8U, the AF488 count increased with the increase of, and reached a maximum value when the amount of Endo IV reached 4U. Therefore, 4U of Endo IV was used for subsequent studies. As shown in Figure 8, when the amount of Endo IV was fixed at 4U, the AF488 count increased with the increase of, and reached a maximum value when the amount of TdT reached 6U. Therefore, 6U of TdT was used in subsequent studies.
7.基因组DNA分离的端粒含量的检测7. Detection of Telomere Content in Genomic DNA Isolation
为了收集基因组DNA端粒,将端粒特异性3'-生物素化的捕获探针和消化的基因组DNA片段在95℃加热5分钟,然后缓慢冷却至室温,通过磁珠分离生物素化的捕获探针,再将磁珠与200mM NaOH在室温下孵育30分钟以释放端粒链,最后将具有端粒链的上清液与磁珠分离,并通过NanoDrop 2000c分光光度计测量端粒量。To collect genomic DNA telomeres, telomere-specific 3'-biotinylated capture probes and digested genomic DNA fragments were heated at 95 °C for 5 min and then slowly cooled to room temperature. The biotinylated capture probes were separated by magnetic beads, and the magnetic beads were incubated with 200 mM NaOH at room temperature for 30 min to release the telomere chains. Finally, the supernatant with the telomere chains was separated from the magnetic beads, and the telomere amount was measured by NanoDrop 2000c spectrophotometer.
8.计算端粒中氧化损伤个数8. Calculate the number of oxidative damage in telomeres
在该方法中,所有游离的3'羟基(3'-OH)仅由切除的氧化损伤位点产生,并且它们可以诱导TdT介导的聚合延伸从而启动Endo IV辅助环状切割信号探针以释放丰富的AF488荧光团。测得的AF488计数与氧化损伤水平成正比。氧化碱损伤,尤其是8-oxoG和FapyG是最常见的氧化性DNA损伤。因此,在的研究中,通过稀释已知浓度的含有两个8-oxoG碱基(Telo-2)的合成端粒来建立标准曲线,得到的对数线性回归方程N=1632.6+101.16log10C,其中N是测量的AF488计数,C是Telo-2的浓度(图4A)。进一步测量了从不同量的基因组DNA中分离的端粒中的氧化损伤(图5B)。在0.1到100ng范围内,AF488计数随着基因组DNA数量的增加而提高。标准曲线用于计算人端粒的绝对氧化损伤水平。每个细胞中人端粒的Fpg敏感性氧化损伤位点的数量计算如下:In this method, all free 3'hydroxyl groups (3'-OH) are generated only by the excised oxidative damage sites, and they can induce TdT-mediated polymerization extension to initiate Endo IV-assisted circular cleavage of the signal probe to release abundant AF488 fluorophores. The measured AF488 counts are proportional to the level of oxidative damage. Oxidative base damage, especially 8-oxoG and FapyG, are the most common oxidative DNA damage. Therefore, in the study, a standard curve was established by diluting synthetic telomeres containing two 8-oxoG bases (Telo-2) of known concentrations, and the resulting logarithmic linear regression equation was N=1632.6+101.16log10 C, where N is the measured AF488 count and C is the concentration of Telo-2 (Figure 4A). Oxidative damage in telomeres isolated from different amounts of genomic DNA was further measured (Figure 5B). In the range of 0.1 to 100 ng, AF488 counts increased with the increase in the amount of genomic DNA. The standard curve was used to calculate the absolute oxidative damage level of human telomeres. The number of Fpg-sensitive oxidative damage sites per cell at human telomeres was calculated as follows:
(1)将从不同量的基因组DNA分离的人端粒中获得的AF488计数(图5B)分别代入标准曲线N=1632.6+101.16log10C。因此,回归方程是:获得的AF488计数=1632.6+101.16log10C并且可以计算浓度C。(1) The AF488 counts obtained from human telomeres isolated from different amounts of genomic DNA ( FIG. 5B ) were respectively substituted into the standard curve N=1632.6+101.16log10 C. Therefore, the regression equation is: AF488 counts obtained=1632.6+101.16log10 C and the concentration C can be calculated.
(2)从不同基因组DNA分离的人端粒中对Fpg敏感的氧化损伤位点(N-telo)的数量为:N-telo=计算浓度C×2(标准曲线中的Telo-2含有两个8-oxoG碱基)×反应体积(10μL)×阿伏加德罗常数=计算浓度C(mol/L)×2×10×10-6L×6.02×1023。(2) The number of Fpg-sensitive oxidative damage sites (N-telo) in human telomeres isolated from different genomic DNAs is: N-telo = calculated concentration C × 2 (Telo-2 in the standard curve contains two 8-oxoG bases) × reaction volume (10 μL) × Avogadro constant = calculated concentration C (mol/L) × 2 × 10 × 10-6 L × 6.02 × 1023 .
(3)每个细胞中人端粒的Fpg敏感性氧化损伤位点的数量计算如下:计算的N-telo/细胞数对应的基因组DNA量。(3) The number of Fpg-sensitive oxidative damage sites on human telomeres per cell was calculated as follows: the amount of genomic DNA corresponding to the calculated number of N-telo/cell.
因此,估计每个细胞端粒中的氧化损伤的数量为每个HeLa细胞34-44(表2)。Therefore, the number of oxidative lesions in each cell telomere was estimated to be 34-44 per HeLa cell (Table 2).
9.检测不同人细胞系端粒的氧化损伤9. Detection of oxidative damage in telomeres of different human cell lines
该方法测量了不同人细胞系中端粒的氧化损伤水平,包括人肺腺癌细胞系(A549细胞),人结肠癌细胞系(SW480细胞)和正常人肝细胞系(HL-7702细胞)。如图10所示,观察到端粒中不同的氧化鸟嘌呤损伤水平,癌细胞(即HeLa细胞,A549细胞和SW480细胞)中的氧化鸟嘌呤损伤水平远高于正常细胞(即HL-7702细胞),与癌症患者的氧化损伤一致。This method measured the oxidative damage levels of telomeres in different human cell lines, including human lung adenocarcinoma cell line (A549 cells), human colon cancer cell line (SW480 cells) and normal human liver cell line (HL-7702 cells). As shown in Figure 10, different levels of oxidative guanine damage in telomeres were observed, and the levels of oxidative guanine damage in cancer cells (i.e., HeLa cells, A549 cells and SW480 cells) were much higher than those in normal cells (i.e., HL-7702 cells), which is consistent with oxidative damage in cancer patients.
应注意的是,以上实例仅用于说明本发明的技术方案而非对其进行限制。尽管参照所给出的实例对本发明进行了详细说明,但是本领域的普通技术人员可根据需要对本发明的技术方案进行修改或者等同替换,而不脱离本发明技术方案的精神和范围。It should be noted that the above examples are only used to illustrate the technical solution of the present invention rather than to limit it. Although the present invention is described in detail with reference to the given examples, those skilled in the art may modify or replace the technical solution of the present invention as needed without departing from the spirit and scope of the technical solution of the present invention.
SEQUENCE LISTINGSEQUENCE LISTING
<110> 山东师范大学<110> Shandong Normal University
<120> 一种检测端粒内氧化性损伤的荧光化学传感器及其检测方法和应用<120> A fluorescent chemical sensor for detecting oxidative damage in telomeres and its detection method and application
<130><130>
<160> 9<160> 9
<170> PatentIn version 3.3<170> PatentIn version 3.3
<210> 1<210> 1
<211> 24<211> 24
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 1<400> 1
ttagggttag ggttagggtt aggg 24ttagggttag ggttagggtt aggg 24
<210> 2<210> 2
<211> 22<211> 22
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 2<400> 2
ttagggttag ggttagggtt addc 22ttagggttag ggttagggtt addc 22
<210> 3<210> 3
<211> 24<211> 24
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 3<400> 3
ttagggttag ggttagggtt aggg 24ttagggttag ggttagggtt aggg 24
<210> 4<210> 4
<211> 21<211> 21
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 4<400> 4
ttagggttag ggttagggtt a 21ttagggttag ggttagggtt a 21
<210> 5<210> 5
<211> 18<211> 18
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 5<400> 5
cctaacccta accctaaddc 18cctaacccta accctaaddc 18
<210> 6<210> 6
<211> 24<211> 24
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 6<400> 6
cctaacccta accctaaccc tttt 24cctaacccta accctaaccc tttt 24
<210> 7<210> 7
<211> 18<211> 18
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 7<400> 7
tttttttttx tttttttt 18tttttttttx ttttttttt 18
<210> 8<210> 8
<211> 24<211> 24
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 8<400> 8
gtagttggag ctggtggcgt aggc 24gtagttggag ctggtggcgt aggc 24
<210> 9<210> 9
<211> 24<211> 24
<212> DNA<212> DNA
<213> 人工合成<213> Synthesis
<400> 9<400> 9
atgcaacgga caagaagatc agta 24atgcaacgga caagaagatc agta 24
Claims (16)
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201910478044.8ACN110144384B (en) | 2019-06-03 | 2019-06-03 | Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereof |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201910478044.8ACN110144384B (en) | 2019-06-03 | 2019-06-03 | Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereof |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN110144384A CN110144384A (en) | 2019-08-20 |
| CN110144384Btrue CN110144384B (en) | 2023-03-24 |
Family
ID=67590034
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201910478044.8AActiveCN110144384B (en) | 2019-06-03 | 2019-06-03 | Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereof |
Country Status (1)
| Country | Link |
|---|---|
| CN (1) | CN110144384B (en) |
Families Citing this family (8)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN111778316B (en)* | 2020-06-09 | 2022-12-27 | 山东师范大学 | Fluorescent probe based on oxidative damage basic group, kit and method for directly detecting DNA methylation |
| CN112852922B (en)* | 2021-01-14 | 2022-12-27 | 山东师范大学 | Fluorescent biosensor for detecting DNA methylation, detection method and application |
| CN112662777B (en)* | 2021-01-19 | 2022-06-24 | 山东师范大学 | Compound and method for simultaneously detecting multiple long-chain non-coding RNAs |
| CN113049559A (en)* | 2021-03-23 | 2021-06-29 | 华中科技大学 | Sperm DNA fragmentation fluorescence detection method, detection kit and application thereof |
| CN113652473B (en)* | 2021-06-30 | 2023-05-05 | 山东师范大学 | Fluorescent chemical sensor for detecting DNA damage site by single molecule, method and application |
| CN113684244A (en)* | 2021-08-19 | 2021-11-23 | 青岛科技大学 | A Label-Free, Homogeneous Bioluminescence Detection Method for DNA Damage |
| CN114507706B (en)* | 2022-01-14 | 2024-04-05 | 山东师范大学 | Biosensor based on enzymatic DNA repair cascade driven fluorophore encoding/decoding and its application in telomerase detection |
| CN114507713A (en)* | 2022-01-14 | 2022-05-17 | 山东师范大学 | Bsu polymerase mediated fluorescence coding sensor and application thereof in 8-oxo-7, 8-dihydroguanine detection |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN106939336A (en)* | 2017-03-14 | 2017-07-11 | 国家体育总局体育科学研究所 | The method for detecting oxidative damage DNA |
| CN107760762A (en)* | 2017-09-30 | 2018-03-06 | 山东师范大学 | A kind of fluorescence chemical sensor and its detection method of detection DNA adenine methyltransferases |
| CN108359448A (en)* | 2018-02-05 | 2018-08-03 | 天津大学 | Three dendritic nucleic acid nano silver fluorescence clusters of one kind and Preparation method and use |
| CN109161579A (en)* | 2018-09-17 | 2019-01-08 | 山东师范大学 | Constant-temperature amplification method based on ligase and its application in polynucleotide kinase detection |
Family Cites Families (1)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| TW201531569A (en)* | 2014-02-12 | 2015-08-16 | Taiwan Sugar Corp | Use of double-stranded nucleic acid molecule and S1 nuclease for detecting the activity of nucleic acid repair enzyme |
- 2019
- 2019-06-03CNCN201910478044.8Apatent/CN110144384B/enactiveActive
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN106939336A (en)* | 2017-03-14 | 2017-07-11 | 国家体育总局体育科学研究所 | The method for detecting oxidative damage DNA |
| CN107760762A (en)* | 2017-09-30 | 2018-03-06 | 山东师范大学 | A kind of fluorescence chemical sensor and its detection method of detection DNA adenine methyltransferases |
| CN108359448A (en)* | 2018-02-05 | 2018-08-03 | 天津大学 | Three dendritic nucleic acid nano silver fluorescence clusters of one kind and Preparation method and use |
| CN109161579A (en)* | 2018-09-17 | 2019-01-08 | 山东师范大学 | Constant-temperature amplification method based on ligase and its application in polynucleotide kinase detection |
Non-Patent Citations (2)
| Title |
|---|
| DNA修复酶活性检测的荧光生物传感方法研究;吴玉姝;《中国博士学位全文数据库 工程科技I辑》;20161115(第11期);第4.4节* |
| 单分子水平检测人类端粒内氧化性损伤;华若男;《中国优秀硕士学位论文全文数据库 医药卫生科技辑》;20200815(第8期);E059-52* |
Also Published As
| Publication number | Publication date |
|---|---|
| CN110144384A (en) | 2019-08-20 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN110144384B (en) | Fluorescent chemical sensor for detecting oxidative damage in telomere as well as detection method and application thereof | |
| JP6571895B1 (en) | Nucleic acid probe and genomic fragment detection method | |
| Li et al. | Rolling circle amplification-driven encoding of different fluorescent molecules for simultaneous detection of multiple DNA repair enzymes at the single-molecule level | |
| Degliangeli et al. | Nanotechnology‐based strategies for the detection and quantification of MicroRNA | |
| CN111154839B (en) | A fluorescent chemical sensor for simultaneous detection of multiple DNA glycosylases, its detection method and application | |
| Wang et al. | A copper-free and enzyme-free click chemistry-mediated single quantum dot nanosensor for accurate detection of microRNAs in cancer cells and tissues | |
| CN112852922B (en) | Fluorescent biosensor for detecting DNA methylation, detection method and application | |
| CN112725416B (en) | Fluorescence sensor for simultaneously detecting hOGG1 and hAAG, and detection method and application thereof | |
| CN110484602B (en) | Fluorescent chemical sensor for detecting DNA damage in genome by single molecule counting, method and application | |
| CN111778316B (en) | Fluorescent probe based on oxidative damage basic group, kit and method for directly detecting DNA methylation | |
| WO2023164694A2 (en) | ONE-POT ENDONUCLEOLYTICALLY EXPONENTIATED ROLLING CIRCLE AMPLIFICATION BY CRISPR-CAS12a | |
| CN113088557B (en) | Fluorescent chemical sensor for simultaneous detection of multiple DNA glycosylases, detection method and application thereof | |
| Zhang et al. | Detection of DNA 3'-phosphatase activity based on exonuclease III-assisted cascade recycling amplification reaction | |
| CN109251960B (en) | Method for detecting Dam methyltransferase activity based on base excision repair induction | |
| CN113755557A (en) | Single quantum dot-based multi-primer generation rolling circle amplification nanosensor and single nucleotide polymorphism detection method | |
| CN107083437B (en) | A method for the ultrasensitive simultaneous detection of multiple DNA glycosylases using inherently fluorescent nucleotides | |
| CN114410793B (en) | Method for detecting FEN1 activity by label-free fluorescence | |
| Al-Hawary et al. | Isothermal amplification methods in cancer-related miRNA detection; a new paradigm in study of cancer pathology | |
| JP2019512214A (en) | Single stranded nucleic acid for detecting nucleic acid or protein in real time and detection method using the same | |
| CN114921526B (en) | Methylation-driven construction of dual-color luminescent RNA nanosensors for label-free and ultrasensitive detection of multiple DNA methyltransferases | |
| CN114507706B (en) | Biosensor based on enzymatic DNA repair cascade driven fluorophore encoding/decoding and its application in telomerase detection | |
| CN113652473B (en) | Fluorescent chemical sensor for detecting DNA damage site by single molecule, method and application | |
| CN114480613B (en) | A detection method for MazF-mediated FTO enzyme and an inhibitor screening method | |
| CN116716395A (en) | Preparation method of a new biosensing platform for self-primed amplification of tandem Cas12a and [Cu(tz)] | |
| Zhang et al. | Single-molecule counting of oxidative DNA damage in telomeres from cancer cells |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| GR01 | Patent grant | ||
| GR01 | Patent grant |