CN110028564B - Rice salt tolerance transcription factor OsC3H38 and its encoding gene and application - Google Patents
Rice salt tolerance transcription factor OsC3H38 and its encoding gene and applicationDownload PDFInfo
- Publication number
- CN110028564B CN110028564BCN201910219494.5ACN201910219494ACN110028564BCN 110028564 BCN110028564 BCN 110028564BCN 201910219494 ACN201910219494 ACN 201910219494ACN 110028564 BCN110028564 BCN 110028564B
- Authority
- CN
- China
- Prior art keywords
- osc3h38
- rice
- salt
- arg
- tolerance
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Expired - Fee Related
Links
- 101000658079Oryza sativa subsp. japonica Splicing factor U2af small subunit BProteins0.000titleclaimsabstractdescription76
- 235000007164Oryza sativaNutrition0.000titleclaimsabstractdescription74
- 235000009566riceNutrition0.000titleclaimsabstractdescription71
- 230000015784hyperosmotic salinity responseEffects0.000titleclaimsabstractdescription28
- 108090000623proteins and genesProteins0.000titleabstractdescription26
- 108091023040Transcription factorProteins0.000titleabstractdescription16
- 102000040945Transcription factorHuman genes0.000titleabstractdescription16
- 240000007594Oryza sativaSpecies0.000titledescription4
- 241000209094OryzaSpecies0.000claimsabstractdescription71
- 239000002773nucleotideSubstances0.000claimsabstractdescription3
- 125000003729nucleotide groupChemical group0.000claimsabstractdescription3
- 238000000034methodMethods0.000claimsdescription6
- 230000009261transgenic effectEffects0.000abstractdescription44
- 230000014509gene expressionEffects0.000abstractdescription32
- 230000002018overexpressionEffects0.000abstractdescription11
- 239000003550markerSubstances0.000abstractdescription6
- 238000011161developmentMethods0.000abstractdescription3
- 125000003275alpha amino acid groupChemical group0.000abstract1
- 230000035882stressEffects0.000description19
- 230000001105regulatory effectEffects0.000description15
- 150000003839saltsChemical class0.000description13
- FAPWRFPIFSIZLT-UHFFFAOYSA-MSodium chlorideChemical compound[Na+].[Cl-]FAPWRFPIFSIZLT-UHFFFAOYSA-M0.000description12
- 238000011282treatmentMethods0.000description12
- 240000004808Saccharomyces cerevisiaeSpecies0.000description10
- 241000196324EmbryophytaSpecies0.000description8
- 101710185494Zinc finger proteinProteins0.000description6
- 102100023597Zinc finger protein 816Human genes0.000description6
- 230000000694effectsEffects0.000description6
- 239000000463materialSubstances0.000description6
- 239000011780sodium chlorideSubstances0.000description6
- 108091030071RNAIProteins0.000description5
- 230000009368gene silencing by RNAEffects0.000description5
- 239000002609mediumSubstances0.000description5
- 230000029279positive regulation of transcription, DNA-dependentEffects0.000description5
- 239000013598vectorSubstances0.000description5
- 108091032973(ribonucleotides)n+mProteins0.000description4
- 108020004414DNAProteins0.000description4
- ONIBWKKTOPOVIA-BYPYZUCNSA-NL-ProlineChemical compoundOC(=O)[C@@H]1CCCN1ONIBWKKTOPOVIA-BYPYZUCNSA-N0.000description4
- ONIBWKKTOPOVIA-UHFFFAOYSA-NProlineNatural productsOC(=O)C1CCCN1ONIBWKKTOPOVIA-UHFFFAOYSA-N0.000description4
- 210000004027cellAnatomy0.000description4
- 229930002875chlorophyllNatural products0.000description4
- 235000019804chlorophyllNutrition0.000description4
- ATNHDLDRLWWWCB-AENOIHSZSA-Mchlorophyll aChemical compoundC1([C@@H](C(=O)OC)C(=O)C2=C3C)=C2N2C3=CC(C(CC)=C3C)=[N+]4C3=CC3=C(C=C)C(C)=C5N3[Mg-2]42[N+]2=C1[C@@H](CCC(=O)OC\C=C(/C)CCC[C@H](C)CCC[C@H](C)CCCC(C)C)[C@H](C)C2=C5ATNHDLDRLWWWCB-AENOIHSZSA-M0.000description4
- 230000012010growthEffects0.000description4
- 230000001965increasing effectEffects0.000description4
- 241000589158AgrobacteriumSpecies0.000description3
- WSMYVTOQOOLQHP-UHFFFAOYSA-NMalondialdehydeChemical compoundO=CCC=OWSMYVTOQOOLQHP-UHFFFAOYSA-N0.000description3
- 230000036579abiotic stressEffects0.000description3
- 238000004458analytical methodMethods0.000description3
- 230000009286beneficial effectEffects0.000description3
- 238000010367cloningMethods0.000description3
- 239000013604expression vectorSubstances0.000description3
- 229940118019malondialdehydeDrugs0.000description3
- XJFPXLWGZWAWRQ-UHFFFAOYSA-N2-[[2-[[2-[[2-[[2-[(2-azaniumylacetyl)amino]acetyl]amino]acetyl]amino]acetyl]amino]acetyl]amino]acetateChemical compoundNCC(=O)NCC(=O)NCC(=O)NCC(=O)NCC(=O)NCC(O)=OXJFPXLWGZWAWRQ-UHFFFAOYSA-N0.000description2
- OPIFSICVWOWJMJ-AEOCFKNESA-N5-bromo-4-chloro-3-indolyl beta-D-galactosideChemical compoundO[C@@H]1[C@@H](O)[C@@H](O)[C@@H](CO)O[C@H]1OC1=CNC2=CC=C(Br)C(Cl)=C12OPIFSICVWOWJMJ-AEOCFKNESA-N0.000description2
- CYXCAHZVPFREJD-LURJTMIESA-NArg-Gly-GlyChemical compoundNC(=N)NCCC[C@H](N)C(=O)NCC(=O)NCC(O)=OCYXCAHZVPFREJD-LURJTMIESA-N0.000description2
- PGUYEUCYVNZGGV-QWRGUYRKSA-NAsp-Gly-TyrChemical compoundOC(=O)C[C@H](N)C(=O)NCC(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1PGUYEUCYVNZGGV-QWRGUYRKSA-N0.000description2
- IJGRMHOSHXDMSA-UHFFFAOYSA-NAtomic nitrogenChemical compoundN#NIJGRMHOSHXDMSA-UHFFFAOYSA-N0.000description2
- 241000620209Escherichia coli DH5[alpha]Species0.000description2
- 125000002707L-tryptophyl groupChemical group[H]C1=C([H])C([H])=C2C(C([C@](N([H])[H])(C(=O)[*])[H])([H])[H])=C([H])N([H])C2=C1[H]0.000description2
- 244000061176Nicotiana tabacumSpecies0.000description2
- 235000002637Nicotiana tabacumNutrition0.000description2
- 238000011529RT qPCRMethods0.000description2
- 108700008625Reporter GenesProteins0.000description2
- 108010068380arginylarginineProteins0.000description2
- 238000001514detection methodMethods0.000description2
- 108010063718gamma-glutamylaspartic acidProteins0.000description2
- 230000035784germinationEffects0.000description2
- 239000013612plasmidSubstances0.000description2
- 102000004169proteins and genesHuman genes0.000description2
- 238000003259recombinant expressionMethods0.000description2
- 238000012163sequencing techniqueMethods0.000description2
- 230000009466transformationEffects0.000description2
- 102000007469ActinsHuman genes0.000description1
- 108010085238ActinsProteins0.000description1
- JYEBJTDTPNKQJG-FXQIFTODSA-NAla-Asn-MetChemical compoundC[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCSC)C(=O)O)NJYEBJTDTPNKQJG-FXQIFTODSA-N0.000description1
- WJRXVTCKASUIFF-FXQIFTODSA-NAla-Cys-ArgChemical compound[H]N[C@@H](C)C(=O)N[C@@H](CS)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OWJRXVTCKASUIFF-FXQIFTODSA-N0.000description1
- 241000219194ArabidopsisSpecies0.000description1
- SGYSTDWPNPKJPP-GUBZILKMSA-NArg-Ala-ArgChemical compoundNC(=N)NCCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OSGYSTDWPNPKJPP-GUBZILKMSA-N0.000description1
- VBFJESQBIWCWRL-DCAQKATOSA-NArg-Ala-LysChemical compoundNCCCC[C@@H](C(O)=O)NC(=O)[C@H](C)NC(=O)[C@@H](N)CCCNC(N)=NVBFJESQBIWCWRL-DCAQKATOSA-N0.000description1
- JGDGLDNAQJJGJI-AVGNSLFASA-NArg-Arg-HisChemical compoundC1=C(NC=N1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCN=C(N)N)NC(=O)[C@H](CCCN=C(N)N)NJGDGLDNAQJJGJI-AVGNSLFASA-N0.000description1
- HJVGMOYJDDXLMI-AVGNSLFASA-NArg-Arg-LysChemical compoundNCCCC[C@@H](C(O)=O)NC(=O)[C@H](CCCNC(N)=N)NC(=O)[C@@H](N)CCCNC(N)=NHJVGMOYJDDXLMI-AVGNSLFASA-N0.000description1
- UISQLSIBJKEJSS-GUBZILKMSA-NArg-Arg-SerChemical compoundNC(N)=NCCC[C@H](N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)N[C@@H](CO)C(O)=OUISQLSIBJKEJSS-GUBZILKMSA-N0.000description1
- UFBURHXMKFQVLM-CIUDSAMLSA-NArg-Glu-SerChemical compound[H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CO)C(O)=OUFBURHXMKFQVLM-CIUDSAMLSA-N0.000description1
- PNIGSVZJNVUVJA-BQBZGAKWSA-NArg-Gly-AsnChemical compoundNC(N)=NCCC[C@H](N)C(=O)NCC(=O)N[C@@H](CC(N)=O)C(O)=OPNIGSVZJNVUVJA-BQBZGAKWSA-N0.000description1
- YBIAYFFIVAZXPK-AVGNSLFASA-NArg-His-ArgChemical compound[H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OYBIAYFFIVAZXPK-AVGNSLFASA-N0.000description1
- UIUXXFIKWQVMEX-UFYCRDLUSA-NArg-Phe-TyrChemical compound[H]N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=OUIUXXFIKWQVMEX-UFYCRDLUSA-N0.000description1
- AWMAZIIEFPFHCP-RCWTZXSCSA-NArg-Pro-ThrChemical compound[H]N[C@@H](CCCNC(N)=N)C(=O)N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)O)C(O)=OAWMAZIIEFPFHCP-RCWTZXSCSA-N0.000description1
- NKTLGLBAGUJEGA-BIIVOSGPSA-NAsn-Cys-ProChemical compoundC1C[C@@H](N(C1)C(=O)[C@H](CS)NC(=O)[C@H](CC(=O)N)N)C(=O)ONKTLGLBAGUJEGA-BIIVOSGPSA-N0.000description1
- PNHQRQTVBRDIEF-CIUDSAMLSA-NAsn-Leu-AlaChemical compoundC[C@@H](C(=O)O)NC(=O)[C@H](CC(C)C)NC(=O)[C@H](CC(=O)N)NPNHQRQTVBRDIEF-CIUDSAMLSA-N0.000description1
- LNENWJXDHCFVOF-DCAQKATOSA-NAsp-His-MetChemical compoundCSCC[C@@H](C(=O)O)NC(=O)[C@H](CC1=CN=CN1)NC(=O)[C@H](CC(=O)O)NLNENWJXDHCFVOF-DCAQKATOSA-N0.000description1
- RTXQQDVBACBSCW-CFMVVWHZSA-NAsp-Ile-TyrChemical compound[H]N[C@@H](CC(O)=O)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(O)=ORTXQQDVBACBSCW-CFMVVWHZSA-N0.000description1
- LIJXJYGRSRWLCJ-IHRRRGAJSA-NAsp-Phe-ArgChemical compound[H]N[C@@H](CC(O)=O)C(=O)N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OLIJXJYGRSRWLCJ-IHRRRGAJSA-N0.000description1
- 241000894006BacteriaSpecies0.000description1
- BUIYOWKUSCTBRE-CIUDSAMLSA-NCys-Arg-GlnChemical compound[H]N[C@@H](CS)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(N)=O)C(O)=OBUIYOWKUSCTBRE-CIUDSAMLSA-N0.000description1
- BCFXQBXXDSEHRS-FXQIFTODSA-NCys-Ser-ArgChemical compound[H]N[C@@H](CS)C(=O)N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OBCFXQBXXDSEHRS-FXQIFTODSA-N0.000description1
- CLPQUWHBWXFJOX-BQBZGAKWSA-NGln-Gly-GlnChemical compound[H]N[C@@H](CCC(N)=O)C(=O)NCC(=O)N[C@@H](CCC(N)=O)C(O)=OCLPQUWHBWXFJOX-BQBZGAKWSA-N0.000description1
- FYBSCGZLICNOBA-XQXXSGGOSA-NGlu-Ala-ThrChemical compound[H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](C)C(=O)N[C@@H]([C@@H](C)O)C(O)=OFYBSCGZLICNOBA-XQXXSGGOSA-N0.000description1
- CGYDXNKRIMJMLV-GUBZILKMSA-NGlu-Arg-GluChemical compound[H]N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=OCGYDXNKRIMJMLV-GUBZILKMSA-N0.000description1
- MUSGDMDGNGXULI-DCAQKATOSA-NGlu-Glu-LeuChemical compoundCC(C)C[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CCC(O)=OMUSGDMDGNGXULI-DCAQKATOSA-N0.000description1
- FMBWLLMUPXTXFC-SDDRHHMPSA-NGlu-Lys-ProChemical compoundC1C[C@@H](N(C1)C(=O)[C@H](CCCCN)NC(=O)[C@H](CCC(=O)O)N)C(=O)OFMBWLLMUPXTXFC-SDDRHHMPSA-N0.000description1
- UPOJUWHGMDJUQZ-IUCAKERBSA-NGly-Arg-ArgChemical compoundNC(=N)NCCC[C@H](NC(=O)CN)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OUPOJUWHGMDJUQZ-IUCAKERBSA-N0.000description1
- OGCIHJPYKVSMTE-YUMQZZPRSA-NGly-Arg-GluChemical compound[H]NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CCC(O)=O)C(O)=OOGCIHJPYKVSMTE-YUMQZZPRSA-N0.000description1
- KKBWDNZXYLGJEY-UHFFFAOYSA-NGly-Arg-ProNatural productsNCC(=O)NC(CCNC(=N)N)C(=O)N1CCCC1C(=O)OKKBWDNZXYLGJEY-UHFFFAOYSA-N0.000description1
- GWCRIHNSVMOBEQ-BQBZGAKWSA-NGly-Arg-SerChemical compound[H]NCC(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=OGWCRIHNSVMOBEQ-BQBZGAKWSA-N0.000description1
- JSNNHGHYGYMVCK-XVKPBYJWSA-NGly-Glu-ValChemical compound[H]NCC(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](C(C)C)C(O)=OJSNNHGHYGYMVCK-XVKPBYJWSA-N0.000description1
- HQRHFUYMGCHHJS-LURJTMIESA-NGly-Gly-ArgChemical compoundNCC(=O)NCC(=O)N[C@H](C(O)=O)CCCN=C(N)NHQRHFUYMGCHHJS-LURJTMIESA-N0.000description1
- UFPXDFOYHVEIPI-BYPYZUCNSA-NGly-Gly-AspChemical compoundNCC(=O)NCC(=O)N[C@H](C(O)=O)CC(O)=OUFPXDFOYHVEIPI-BYPYZUCNSA-N0.000description1
- YOBGUCWZPXJHTN-BQBZGAKWSA-NGly-Ser-ArgChemical compoundNCC(=O)N[C@@H](CO)C(=O)N[C@H](C(O)=O)CCCN=C(N)NYOBGUCWZPXJHTN-BQBZGAKWSA-N0.000description1
- DBUNZBWUWCIELX-JHEQGTHGSA-NGly-Thr-GluChemical compound[H]NCC(=O)N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CCC(O)=O)C(O)=ODBUNZBWUWCIELX-JHEQGTHGSA-N0.000description1
- DNVDEMWIYLVIQU-RCOVLWMOSA-NGly-Val-AspChemical compoundNCC(=O)N[C@@H](C(C)C)C(=O)N[C@@H](CC(O)=O)C(O)=ODNVDEMWIYLVIQU-RCOVLWMOSA-N0.000description1
- 244000068988Glycine maxSpecies0.000description1
- 235000010469Glycine maxNutrition0.000description1
- PYNUBZSXKQKAHL-UWVGGRQHSA-NHis-Gly-ArgChemical compound[H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=OPYNUBZSXKQKAHL-UWVGGRQHSA-N0.000description1
- OEROYDLRVAYIMQ-YUMQZZPRSA-NHis-Gly-AspChemical compound[H]N[C@@H](CC1=CNC=N1)C(=O)NCC(=O)N[C@@H](CC(O)=O)C(O)=OOEROYDLRVAYIMQ-YUMQZZPRSA-N0.000description1
- VYUXYMRNGALHEA-DLOVCJGASA-NHis-Leu-AlaChemical compound[H]N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](C)C(O)=OVYUXYMRNGALHEA-DLOVCJGASA-N0.000description1
- YAEKRYQASVCDLK-JYJNAYRXSA-NHis-Phe-GluChemical compoundC1=CC=C(C=C1)C[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)O)NC(=O)[C@H](CC2=CN=CN2)NYAEKRYQASVCDLK-JYJNAYRXSA-N0.000description1
- LGMUPVWZEYYUMU-YVNDNENWSA-NIle-Glu-GlnChemical compoundCC[C@H](C)[C@@H](C(=O)N[C@@H](CCC(=O)O)C(=O)N[C@@H](CCC(=O)N)C(=O)O)NLGMUPVWZEYYUMU-YVNDNENWSA-N0.000description1
- SLQVFYWBGNNOTK-BYULHYEWSA-NIle-Gly-AsnChemical compoundCC[C@H](C)[C@@H](C(=O)NCC(=O)N[C@@H](CC(=O)N)C(=O)O)NSLQVFYWBGNNOTK-BYULHYEWSA-N0.000description1
- ANTFEOSJMAUGIB-KNZXXDILSA-NIle-Thr-ProChemical compoundCC[C@H](C)[C@@H](C(=O)N[C@@H]([C@@H](C)O)C(=O)N1CCC[C@@H]1C(=O)O)NANTFEOSJMAUGIB-KNZXXDILSA-N0.000description1
- FADYJNXDPBKVCA-UHFFFAOYSA-NL-Phenylalanyl-L-lysinNatural productsNCCCCC(C(O)=O)NC(=O)C(N)CC1=CC=CC=C1FADYJNXDPBKVCA-UHFFFAOYSA-N0.000description1
- DPWGZWUMUUJQDT-IUCAKERBSA-NLeu-Gln-GlyChemical compoundCC(C)C[C@H](N)C(=O)N[C@@H](CCC(N)=O)C(=O)NCC(O)=ODPWGZWUMUUJQDT-IUCAKERBSA-N0.000description1
- BTNXKBVLWJBTNR-SRVKXCTJSA-NLeu-His-AsnChemical compound[H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CNC=N1)C(=O)N[C@@H](CC(N)=O)C(O)=OBTNXKBVLWJBTNR-SRVKXCTJSA-N0.000description1
- WFCKERTZVCQXKH-KBPBESRZSA-NLeu-Tyr-GlyChemical compound[H]N[C@@H](CC(C)C)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)NCC(O)=OWFCKERTZVCQXKH-KBPBESRZSA-N0.000description1
- FLCMXEFCTLXBTL-DCAQKATOSA-NLys-Asp-ArgChemical compoundC(CCN)C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NFLCMXEFCTLXBTL-DCAQKATOSA-N0.000description1
- IVFUVMSKSFSFBT-NHCYSSNCSA-NLys-Ile-GlyChemical compoundOC(=O)CNC(=O)[C@H]([C@@H](C)CC)NC(=O)[C@@H](N)CCCCNIVFUVMSKSFSFBT-NHCYSSNCSA-N0.000description1
- ONGCSGVHCSAATF-CIUDSAMLSA-NMet-Ala-GluChemical compoundCSCC[C@H](N)C(=O)N[C@@H](C)C(=O)N[C@H](C(O)=O)CCC(O)=OONGCSGVHCSAATF-CIUDSAMLSA-N0.000description1
- FWTBMGAKKPSTBT-GUBZILKMSA-NMet-Gln-GluChemical compound[H]N[C@@H](CCSC)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=OFWTBMGAKKPSTBT-GUBZILKMSA-N0.000description1
- 241000699670Mus sp.Species0.000description1
- YBAFDPFAUTYYRW-UHFFFAOYSA-NN-L-alpha-glutamyl-L-leucineNatural productsCC(C)CC(C(O)=O)NC(=O)C(N)CCC(O)=OYBAFDPFAUTYYRW-UHFFFAOYSA-N0.000description1
- PESQCPHRXOFIPX-UHFFFAOYSA-NN-L-methionyl-L-tyrosineNatural productsCSCCC(N)C(=O)NC(C(O)=O)CC1=CC=C(O)C=C1PESQCPHRXOFIPX-UHFFFAOYSA-N0.000description1
- 108010079364N-glycylalanineProteins0.000description1
- KYYMILWEGJYPQZ-IHRRRGAJSA-NPhe-Glu-GluChemical compoundOC(=O)CC[C@@H](C(O)=O)NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H](N)CC1=CC=CC=C1KYYMILWEGJYPQZ-IHRRRGAJSA-N0.000description1
- QTVUPXHPSXZJKH-ULQDDVLXSA-NPhe-Met-HisChemical compoundCSCC[C@@H](C(=O)N[C@@H](CC1=CN=CN1)C(=O)O)NC(=O)[C@H](CC2=CC=CC=C2)NQTVUPXHPSXZJKH-ULQDDVLXSA-N0.000description1
- AGTHXWTYCLLYMC-FHWLQOOXSA-NPhe-Tyr-GluChemical compoundC([C@H](N)C(=O)N[C@@H](CC=1C=CC(O)=CC=1)C(=O)N[C@@H](CCC(O)=O)C(O)=O)C1=CC=CC=C1AGTHXWTYCLLYMC-FHWLQOOXSA-N0.000description1
- FRMKIPSIZSFTTE-HJOGWXRNSA-NPhe-Tyr-PheChemical compound[H]N[C@@H](CC1=CC=CC=C1)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=OFRMKIPSIZSFTTE-HJOGWXRNSA-N0.000description1
- QVIZLAUEAMQKGS-GUBZILKMSA-NPro-Asp-MetChemical compoundCSCC[C@@H](C(=O)O)NC(=O)[C@H](CC(=O)O)NC(=O)[C@@H]1CCCN1QVIZLAUEAMQKGS-GUBZILKMSA-N0.000description1
- VOZIBWWZSBIXQN-SRVKXCTJSA-NPro-Glu-LysChemical compoundNCCCC[C@H](NC(=O)[C@H](CCC(O)=O)NC(=O)[C@@H]1CCCN1)C(O)=OVOZIBWWZSBIXQN-SRVKXCTJSA-N0.000description1
- AQGUSRZKDZYGGV-GMOBBJLQSA-NPro-Ile-AspChemical compound[H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC(O)=O)C(O)=OAQGUSRZKDZYGGV-GMOBBJLQSA-N0.000description1
- LNOWDSPAYBWJOR-PEDHHIEDSA-NPro-Ile-IleChemical compound[H]N1CCC[C@H]1C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H]([C@@H](C)CC)C(O)=OLNOWDSPAYBWJOR-PEDHHIEDSA-N0.000description1
- SNGZLPOXVRTNMB-LPEHRKFASA-NPro-Ser-ProChemical compoundC1C[C@H](NC1)C(=O)N[C@@H](CO)C(=O)N2CCC[C@@H]2C(=O)OSNGZLPOXVRTNMB-LPEHRKFASA-N0.000description1
- YDTUEBLEAVANFH-RCWTZXSCSA-NPro-Val-ThrChemical compoundC[C@@H](O)[C@@H](C(O)=O)NC(=O)[C@H](C(C)C)NC(=O)[C@@H]1CCCN1YDTUEBLEAVANFH-RCWTZXSCSA-N0.000description1
- QVOGDCQNGLBNCR-FXQIFTODSA-NSer-Arg-SerChemical compound[H]N[C@@H](CO)C(=O)N[C@@H](CCCNC(N)=N)C(=O)N[C@@H](CO)C(O)=OQVOGDCQNGLBNCR-FXQIFTODSA-N0.000description1
- ZHYMUFQVKGJNRM-ZLUOBGJFSA-NSer-Cys-AsnChemical compoundOC[C@H](N)C(=O)N[C@@H](CS)C(=O)N[C@H](C(O)=O)CC(N)=OZHYMUFQVKGJNRM-ZLUOBGJFSA-N0.000description1
- UOLGINIHBRIECN-FXQIFTODSA-NSer-Glu-GluChemical compound[H]N[C@@H](CO)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@@H](CCC(O)=O)C(O)=OUOLGINIHBRIECN-FXQIFTODSA-N0.000description1
- AEGUWTFAQQWVLC-BQBZGAKWSA-NSer-Gly-ArgChemical compound[H]N[C@@H](CO)C(=O)NCC(=O)N[C@@H](CCCNC(N)=N)C(O)=OAEGUWTFAQQWVLC-BQBZGAKWSA-N0.000description1
- LWMQRHDTXHQQOV-MXAVVETBSA-NSer-Ile-PheChemical compound[H]N[C@@H](CO)C(=O)N[C@@H]([C@@H](C)CC)C(=O)N[C@@H](CC1=CC=CC=C1)C(O)=OLWMQRHDTXHQQOV-MXAVVETBSA-N0.000description1
- WGDYNRCOQRERLZ-KKUMJFAQSA-NSer-Lys-PheChemical compoundC1=CC=C(C=C1)C[C@@H](C(=O)O)NC(=O)[C@H](CCCCN)NC(=O)[C@H](CO)NWGDYNRCOQRERLZ-KKUMJFAQSA-N0.000description1
- DINQYZRMXGWWTG-GUBZILKMSA-NSer-Pro-ProChemical compoundOC[C@H](N)C(=O)N1CCC[C@H]1C(=O)N1[C@H](C(O)=O)CCC1DINQYZRMXGWWTG-GUBZILKMSA-N0.000description1
- CKDXFSPMIDSMGV-GUBZILKMSA-NSer-Pro-ValChemical compound[H]N[C@@H](CO)C(=O)N1CCC[C@H]1C(=O)N[C@@H](C(C)C)C(O)=OCKDXFSPMIDSMGV-GUBZILKMSA-N0.000description1
- FGBLCMLXHRPVOF-IHRRRGAJSA-NSer-Tyr-ArgChemical compound[H]N[C@@H](CO)C(=O)N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OFGBLCMLXHRPVOF-IHRRRGAJSA-N0.000description1
- XYFISNXATOERFZ-OSUNSFLBSA-NThr-Ile-ValChemical compoundCC[C@H](C)[C@@H](C(=O)N[C@@H](C(C)C)C(=O)O)NC(=O)[C@H]([C@@H](C)O)NXYFISNXATOERFZ-OSUNSFLBSA-N0.000description1
- IMDMLDSVUSMAEJ-HJGDQZAQSA-NThr-Leu-AsnChemical compound[H]N[C@@H]([C@@H](C)O)C(=O)N[C@@H](CC(C)C)C(=O)N[C@@H](CC(N)=O)C(O)=OIMDMLDSVUSMAEJ-HJGDQZAQSA-N0.000description1
- IBBBOLAPFHRDHW-BPUTZDHNSA-NTrp-Asn-ArgChemical compoundC1=CC=C2C(=C1)C(=CN2)C[C@@H](C(=O)N[C@@H](CC(=O)N)C(=O)N[C@@H](CCCN=C(N)N)C(=O)O)NIBBBOLAPFHRDHW-BPUTZDHNSA-N0.000description1
- BEIGSKUPTIFYRZ-SRVKXCTJSA-NTyr-Asp-AspChemical compoundC1=CC(=CC=C1C[C@@H](C(=O)N[C@@H](CC(=O)O)C(=O)N[C@@H](CC(=O)O)C(=O)O)N)OBEIGSKUPTIFYRZ-SRVKXCTJSA-N0.000description1
- XBWKCYFGRXKWGO-SRVKXCTJSA-NTyr-Cys-AsnChemical compound[H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(N)=O)C(O)=OXBWKCYFGRXKWGO-SRVKXCTJSA-N0.000description1
- ARPONUQDNWLXOZ-KKUMJFAQSA-NTyr-Gln-ArgChemical compound[H]N[C@@H](CC1=CC=C(O)C=C1)C(=O)N[C@@H](CCC(N)=O)C(=O)N[C@@H](CCCNC(N)=N)C(O)=OARPONUQDNWLXOZ-KKUMJFAQSA-N0.000description1
- BWVHQINTNLVWGZ-ZKWXMUAHSA-NVal-Cys-AspChemical compoundCC(C)[C@@H](C(=O)N[C@@H](CS)C(=O)N[C@@H](CC(=O)O)C(=O)O)NBWVHQINTNLVWGZ-ZKWXMUAHSA-N0.000description1
- PMXBARDFIAPBGK-DZKIICNBSA-NVal-Glu-TyrChemical compoundCC(C)[C@H](N)C(=O)N[C@@H](CCC(O)=O)C(=O)N[C@H](C(O)=O)CC1=CC=C(O)C=C1PMXBARDFIAPBGK-DZKIICNBSA-N0.000description1
- WBAJDGWKRIHOAC-GVXVVHGQSA-NVal-Lys-GlnChemical compound[H]N[C@@H](C(C)C)C(=O)N[C@@H](CCCCN)C(=O)N[C@@H](CCC(N)=O)C(O)=OWBAJDGWKRIHOAC-GVXVVHGQSA-N0.000description1
- GBIUHAYJGWVNLN-AEJSXWLSSA-NVal-Ser-ProChemical compoundCC(C)[C@@H](C(=O)N[C@@H](CO)C(=O)N1CCC[C@@H]1C(=O)O)NGBIUHAYJGWVNLN-AEJSXWLSSA-N0.000description1
- GBIUHAYJGWVNLN-UHFFFAOYSA-NVal-Ser-ProNatural productsCC(C)C(N)C(=O)NC(CO)C(=O)N1CCCC1C(O)=OGBIUHAYJGWVNLN-UHFFFAOYSA-N0.000description1
- HCHKCACWOHOZIP-UHFFFAOYSA-NZincChemical compound[Zn]HCHKCACWOHOZIP-UHFFFAOYSA-N0.000description1
- KOSRFJWDECSPRO-UHFFFAOYSA-Nalpha-L-glutamyl-L-glutamic acidNatural productsOC(=O)CCC(N)C(=O)NC(CCC(O)=O)C(O)=OKOSRFJWDECSPRO-UHFFFAOYSA-N0.000description1
- 150000001413amino acidsChemical group0.000description1
- 230000003321amplificationEffects0.000description1
- 108010013835arginine glutamateProteins0.000description1
- 108010001271arginyl-glutamyl-arginineProteins0.000description1
- 108010084758arginyl-tyrosyl-aspartic acidProteins0.000description1
- 108010060035arginylprolineProteins0.000description1
- 108010093581aspartyl-prolineProteins0.000description1
- 108010047857aspartylglycineProteins0.000description1
- 230000033228biological regulationEffects0.000description1
- 230000011712cell developmentEffects0.000description1
- 230000024245cell differentiationEffects0.000description1
- 230000010261cell growthEffects0.000description1
- 210000000170cell membraneAnatomy0.000description1
- 230000008859changeEffects0.000description1
- 239000002299complementary DNASubstances0.000description1
- 230000009089cytolysisEffects0.000description1
- 230000002950deficientEffects0.000description1
- 230000018109developmental processEffects0.000description1
- 230000024346drought recoveryEffects0.000description1
- 238000001962electrophoresisMethods0.000description1
- 238000005516engineering processMethods0.000description1
- 210000001339epidermal cellAnatomy0.000description1
- 238000002474experimental methodMethods0.000description1
- 230000004927fusionEffects0.000description1
- 108010055341glutamyl-glutamic acidProteins0.000description1
- VPZXBVLAVMBEQI-UHFFFAOYSA-Nglycyl-DL-alpha-alanineNatural productsOC(=O)C(C)NC(=O)CNVPZXBVLAVMBEQI-UHFFFAOYSA-N0.000description1
- 108010067216glycyl-glycyl-glycineProteins0.000description1
- 108010001064glycyl-glycyl-glycyl-glycineProteins0.000description1
- 108010010096glycyl-glycyl-tyrosineProteins0.000description1
- 108010036413histidylglycineProteins0.000description1
- 230000001939inductive effectEffects0.000description1
- 239000007788liquidSubstances0.000description1
- 238000005259measurementMethods0.000description1
- 239000006870ms-mediumSubstances0.000description1
- 229910052757nitrogenInorganic materials0.000description1
- 238000003199nucleic acid amplification methodMethods0.000description1
- 230000008569processEffects0.000description1
- 108010004914prolylarginineProteins0.000description1
- 108010029020prolylglycineProteins0.000description1
- 238000003753real-time PCRMethods0.000description1
- 238000011084recoveryMethods0.000description1
- 230000008261resistance mechanismEffects0.000description1
- 230000006808response to salt stressEffects0.000description1
- 238000010839reverse transcriptionMethods0.000description1
- 238000003757reverse transcription PCRMethods0.000description1
- 238000012216screeningMethods0.000description1
- 108010026333seryl-prolineProteins0.000description1
- 230000003068static effectEffects0.000description1
- 230000004960subcellular localizationEffects0.000description1
- 230000004083survival effectEffects0.000description1
- 238000012360testing methodMethods0.000description1
- 238000013518transcriptionMethods0.000description1
- 230000035897transcriptionEffects0.000description1
- 230000002103transcriptional effectEffects0.000description1
- 108010017949tyrosyl-glycyl-glycineProteins0.000description1
- 229910021642ultra pure waterInorganic materials0.000description1
- 239000012498ultrapure waterSubstances0.000description1
- 239000011701zincSubstances0.000description1
- 229910052725zincInorganic materials0.000description1
Images








Classifications
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/415—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from plants
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8273—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for drought, cold, salt resistance
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Molecular Biology (AREA)
- Engineering & Computer Science (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Biophysics (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- General Health & Medical Sciences (AREA)
- Biomedical Technology (AREA)
- Biochemistry (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Physics & Mathematics (AREA)
- Microbiology (AREA)
- Plant Pathology (AREA)
- Cell Biology (AREA)
- Botany (AREA)
- Gastroenterology & Hepatology (AREA)
- Medicinal Chemistry (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Measuring Or Testing Involving Enzymes Or Micro-Organisms (AREA)
Abstract
Description
Translated fromChinese技术领域technical field
本发明属于农业生物技术领域,具体涉及水稻耐盐转录因子OsC3H38及其编码基因和应用。The invention belongs to the field of agricultural biotechnology, and particularly relates to a rice salt-tolerant transcription factor OsC3H38 and its encoding gene and application.
背景技术Background technique
转录因子在作物的细胞分化、生长发育、非生物胁迫耐受等过程中发挥重要的作用。锌指蛋白家族是一类具有手指状结构域的转录因子,其通过与靶分子DNA、RNA、DNA-RNA的序列特异性结合,或与其他锌指蛋白结合以及形成同源二聚体结构,在转录和翻译水平上调控基因的表达,锌指蛋白家族是植物中参与非生物胁迫调控的重要转录因子类型。根据锌指蛋白的保守域的结构,锌指蛋白有C2H2型、C3H型、C4型和C6型等很多种类型,其中C3H型是发现较晚的一种锌指蛋白,现有技术对动物中C3H型蛋白的研究较多,而仅研究了水稻、拟南芥、大豆等少数几个植物中的C3H型锌指基因的功能。Transcription factors play an important role in the process of cell differentiation, growth and development, and abiotic stress tolerance of crops. The zinc finger protein family is a class of transcription factors with finger-like domains, which bind to target molecules DNA, RNA, DNA-RNA sequence-specifically, or bind to other zinc finger proteins and form a homodimeric structure, Regulating gene expression at the transcriptional and translational levels, the zinc finger protein family is an important type of transcription factor involved in the regulation of abiotic stress in plants. According to the structure of the conserved domains of zinc finger proteins, there are many types of zinc finger proteins, such as C2H2, C3H, C4 and C6, among which C3H is a late-discovered zinc finger protein. There are many studies on C3H-type proteins, but only the functions of C3H-type zinc finger genes in a few plants such as rice, Arabidopsis, and soybean have been studied.
发明内容SUMMARY OF THE INVENTION
本发明的目的在于提供一种水稻耐盐转录因子OsC3H38。The purpose of the present invention is to provide a rice salt-tolerant transcription factor OsC3H38.
本发明的再一目的在于提供上述水稻耐盐转录因子OsC3H38的编码基因。Another object of the present invention is to provide a gene encoding the above-mentioned rice salt tolerance transcription factor OsC3H38.
本发明的再一目的在于提供含有上述编码基因的重组表达载体。Another object of the present invention is to provide a recombinant expression vector containing the above-mentioned encoding gene.
本发明的再一目的在于提供含有上述编码基因的重组菌株。Another object of the present invention is to provide a recombinant strain containing the above-mentioned encoding gene.
本发明的再一目的在于提供上述水稻耐盐转录因子OsC3H38及其编码基因的应用。Another object of the present invention is to provide the application of the above-mentioned rice salt tolerance transcription factor OsC3H38 and its encoding gene.
本发明的再一目的在于提供提高水稻耐旱性的方法。Another object of the present invention is to provide a method for improving drought tolerance of rice.
本发明的水稻转录因子OsC3H38,其氨基酸序列如SEQ ID No.1所示:The rice transcription factor OsC3H38 of the present invention has an amino acid sequence as shown in SEQ ID No.1:
MAEHLASIFGTEKDRVNCPFYFKIGACRHGDRCSRLHNRPTVSPTIVLANMYQRPDMITPGVDAQGQPIDPEKMQEHFEDFYEDIYEELSKFGEVETLNVCDNLADHMIGNVYVQFREEEQAVAAHNALQGRFYSGRPIIVEYSPVTDFREATCRQFEENSCNRGGYCNFMHVKQIGRELRRKLYGGRSRRSHGRSRSPSPRHRRGNRDRDDFRRERDGYRGGGDGYRGGGGGGGGDGYRGGDSYRGGGGGGRRGGGSRYDRYDDGGRRRHGSPPRRARSPVRESSEERRAKIEQWNREREEKPMAEHLASIFGTEKDRVNCPFYFKIGACRHGDRCSRLHNRPTVSPTIVLANMYQRPDMITPGVDAQGQPIDPEKMQEHFEDFYEDIYEELSKFGEVETLNVCDNLADHMIGNVYVQFREEEQAVAAHNALQGRFYSGRPIIVEYSPVTDFREATCRQFEENSCNRGGYCNFMHVKQIGRELRRKLYGGRSRRSHGRSRSPSPRHRRGNRDRDDFRRERDGYRGGGDGYRGGGGGGGGDGYRGGDSYRGGGGGGRRGGGSRYDRYDDGGRRRHGSPPRRARSPVRESSEERRAKIEQWNREREEKP
本发明的水稻转录因子OsC3H38的编码基因,其核苷酸序列如SEQ ID No.2所示:The nucleotide sequence of the encoding gene of the rice transcription factor OsC3H38 of the present invention is shown in SEQ ID No.2:
ATGGCGGAGCATTTGGCTTCAATATTTGGGACTGAGAAGGATAGGGTTAACTGCCCCTTTTACTTCAAGATTGGGGCTTGCCGTCATGGGGATCGTTGCTCCCGTCTGCACAACAGGCCAACTGTGTCGCCGACAATTGTGCTTGCTAACATGTATCAGCGCCCTGATATGATCACACCAGGAGTCGATGCTCAAGGCCAACCTATTGATCCTGAAAAGATGCAGGAGCACTTTGAGGATTTCTATGAGGATATCTATGAGGAGCTGAGCAAGTTTGGTGAGGTTGAGACCCTCAATGTCTGTGATAACCTTGCTGACCACATGATAGGCAATGTGTATGTCCAGTTCAGGGAGGAAGAACAGGCAGTCGCTGCGCATAATGCCCTTCAAGGTCGCTTCTACTCAGGGCGCCCAATAATTGTTGAGTATTCCCCTGTGACTGATTTCCGTGAGGCGACCTGTAGGCAGTTTGAGGAGAACAGCTGCAACCGTGGGGGCTACTGTAATTTTATGCATGTAAAGCAGATTGGCAGGGAGCTCAGGAGGAAGCTTTATGGGGGGCGATCCAGGAGGAGCCATGGGAGAAGCCGCAGCCCAAGCCCACGCCACAGGAGGGGCAATCGTGACCGTGATGATTTCCGCCGTGAACGTGATGGTTATCGTGGTGGTGGTGATGGTTACCGTGGCGGCGGCGGTGGCGGTGGTGGTGACGGTTACCGCGGCGGTGATAGTTACCGTGGTGGTGGTGGTGGTGGACGCAGGGGTGGGGGCAGCAGGTATGACAGGTATGATGATGGAGGAAGGCGCAGGCATGGCAGCCCCCCAAGGCGTGCAAGGAGCCCAGTCAGGGAAAGCAGCGAGGAACGCAGGGCGAAGATTGAACAGTGGAACCGTGAACGGGAGGAAAAGCCTTGAATGGCGGAGCATTTGGCTTCAATATTTGGGACTGAGAAGGATAGGGTTAACTGCCCCTTTTACTTCAAGATTGGGGCTTGCCGTCATGGGGATCGTTGCTCCCGTCTGCACAACAGGCCAACTGTGTCGCCGACAATTGTGCTTGCTAACATGTATCAGCGCCCTGATATGATCACACCAGGAGTCGATGCTCAAGGCCAACCTATTGATCCTGAAAAGATGCAGGAGCACTTTGAGGATTTCTATGAGGATATCTATGAGGAGCTGAGCAAGTTTGGTGAGGTTGAGACCCTCAATGTCTGTGATAACCTTGCTGACCACATGATAGGCAATGTGTATGTCCAGTTCAGGGAGGAAGAACAGGCAGTCGCTGCGCATAATGCCCTTCAAGGTCGCTTCTACTCAGGGCGCCCAATAATTGTTGAGTATTCCCCTGTGACTGATTTCCGTGAGGCGACCTGTAGGCAGTTTGAGGAGAACAGCTGCAACCGTGGGGGCTACTGTAATTTTATGCATGTAAAGCAGATTGGCAGGGAGCTCAGGAGGAAGCTTTATGGGGGGCGATCCAGGAGGAGCCATGGGAGAAGCCGCAGCCCAAGCCCACGCCACAGGAGGGGCAATCGTGACCGTGATGATTTCCGCCGTGAACGTGATGGTTATCGTGGTGGTGGTGATGGTTACCGTGGCGGCGGCGGTGGCGGTGGTGGTGACGGTTACCGCGGCGGTGATAGTTACCGTGGTGGTGGTGGTGGTGGACGCAGGGGTGGGGGCAGCAGGTATGACAGGTATGATGATGGAGGAAGGCGCAGGCATGGCAGCCCCCCAAGGCGTGCAAGGAGCCCAGTCAGGGAAAGCAGCGAGGAACGCAGGGCGAAGATTGAACAGTGGAACCGTGAACGGGAGGAAAAGCCTTGA
本发明还提供含有上述编码基因的重组表达载体。The present invention also provides a recombinant expression vector containing the above-mentioned encoding gene.
本发明还提供含有上述编码基因的重组菌株。The present invention also provides a recombinant strain containing the above-mentioned encoding gene.
根据本发明的提高水稻耐盐的方法包括向水稻中导入水稻耐盐相关基因OsC3H38的步骤。The method for improving the salt tolerance of rice according to the present invention includes the step of introducing the rice salt tolerance-related gene OsC3H38 into the rice.
本发明的有益效果:Beneficial effects of the present invention:
试验结果表明,在175mM NaCl和5μM ABA处理后,野生型水稻中OsC3H38基因的表达量分别上调5.8倍和8.0倍;OsC3H38蛋白定位在细胞核里,具有转录激活活性,说明OsC3H38是受高盐和ABA诱导表达的转录因子。对经过DNA水平和RNA水平的鉴定的OsC3H38转基因水稻进行耐盐性鉴定,结果发现,OsC3H38基因的上调表达提高了转基因水稻在幼苗期和分蘖期的耐盐性,提高了转基因水稻在盐胁迫条件下的叶绿素含量和脯氨酸含量及胁迫条件下全闭气孔的比率,降低了转基因水稻的丙二醛含量;而OsC3H38基因下调表达对转基因水稻的耐逆表型及耐逆相关生理指标的影响则与OsC3H38基因过表达情况下的相反。OsC3H38基因过表达上调了耐逆标记基因OsLEA3、OsSRO1C和OsRAB21的表达量。因此,表型、生理水平和分子水平均表明OsC3H38基因上调表达提高了转基因植物对高盐的耐受性。The experimental results showed that after 175mM NaCl and 5μM ABA treatment, the expression of OsC3H38 gene in wild-type rice was up-regulated by 5.8 times and 8.0 times, respectively; OsC3H38 protein was located in the nucleus and had transcriptional activation activity, indicating that OsC3H38 was affected by high salt and ABA. Inducible transcription factor. The salt tolerance of the OsC3H38 transgenic rice identified at the DNA level and RNA level was identified. The results showed that the up-regulated expression of the OsC3H38 gene improved the salt tolerance of the transgenic rice at the seedling stage and tillering stage, and improved the transgenic rice under salt stress conditions. The chlorophyll content and proline content and the ratio of fully closed stomata under stress conditions reduced the malondialdehyde content of transgenic rice; while the down-expression of OsC3H38 gene had effects on the stress tolerance phenotype and stress tolerance-related physiological indicators of transgenic rice In contrast to the overexpression of OsC3H38 gene. Overexpression of OsC3H38 up-regulated the expression levels of stress tolerance marker genes OsLEA3, OsSRO1C and OsRAB21. Therefore, the phenotypic, physiological and molecular levels indicated that the up-regulated expression of OsC3H38 increased the tolerance of transgenic plants to high salt.
OsC3H38过表达提高了转基因水稻在不同发育时期的耐盐性,同时改善了耐逆相关的生理指标,提高了耐盐标记基因的表达量。因此,在水稻中提高OsC3H38基因的表达量,可以提高水稻在高盐条件下的存活率,上调其下游耐盐相关基因的表达量,提高了植株的耐盐性,可以用于农作物耐盐品种的培育。Overexpression of OsC3H38 improved the salt tolerance of transgenic rice at different developmental stages, improved physiological indicators related to stress tolerance, and increased the expression of salt tolerance marker genes. Therefore, increasing the expression of the OsC3H38 gene in rice can improve the survival rate of rice under high-salt conditions, up-regulate the expression of its downstream salt-tolerance-related genes, and improve the salt-tolerance of plants, which can be used in crop salt-tolerant varieties 's cultivation.
附图说明Description of drawings
图1显示水稻OsC3H38基因在高盐和ABA处理下的表达量变化;Figure 1 shows the expression changes of rice OsC3H38 gene under high salt and ABA treatments;
图2显示OsC3H38蛋白的亚细胞定位情况;Figure 2 shows the subcellular localization of OsC3H38 protein;
图3显示OSCH38的转录激活活性鉴定结果,其中,A为OsC3H38转基因酵母在不同培养基上的生长情况,B为OsC3H38转基因酵母的X-gal显色结果;Figure 3 shows the identification results of the transcriptional activation activity of OSCH38, wherein A is the growth of OsC3H38 transgenic yeast on different media, and B is the X-gal color development result of OsC3H38 transgenic yeast;
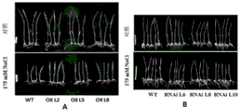
图4显示对获得的OsC3H38转基因水稻的鉴定情况,其中,A为OsC3H38转基因水稻的PCR结果,B为OsC3H38转基因水稻不同株系的基因表达情况,C为OsC3H38RNAi转基因水稻不同株系的基因表达情况;Figure 4 shows the identification of the obtained OsC3H38 transgenic rice, wherein A is the PCR result of OsC3H38 transgenic rice, B is the gene expression of different lines of OsC3H38 transgenic rice, and C is the gene expression of different lines of OsC3H38 RNAi transgenic rice;
图5显示转基因水稻在幼苗期的耐盐性鉴定情况,其中,A为OsC3H38转基因水稻不同株系的生长情况,B为OsC3H38RNAi转基因水稻不同株系的生长情况;Figure 5 shows the identification of salt tolerance of transgenic rice at the seedling stage, wherein A is the growth of different lines of OsC3H38 transgenic rice, and B is the growth of different lines of OsC3H38 RNAi transgenic rice;
图6显示转基因水稻在幼苗期的耐盐性鉴定情况,其中,A为OsC3H38转基因水稻不同株系的地上部分长度,B为OsC3H38RNAi转基因水稻不同株系的地上部分长度;Figure 6 shows the identification of salt tolerance of transgenic rice at the seedling stage, wherein A is the length of the aerial part of different lines of OsC3H38 transgenic rice, and B is the length of the aerial part of different lines of OsC3H38 RNAi transgenic rice;
图7显示对OsC3H38转基因水稻在分蘖期的耐盐性鉴定情况;Figure 7 shows the identification of salt tolerance of OsC3H38 transgenic rice at tillering stage;
图8显示野生型和OsC3H38转基因水稻在盐胁迫前后的不同生理指标的测定结果;Figure 8 shows the measurement results of different physiological indicators of wild-type and OsC3H38 transgenic rice before and after salt stress;
图9显示OsC3H38过表达转基因水稻的耐逆相关标记基因的表达量。Figure 9 shows the expression levels of inverse tolerance-related marker genes in OsC3H38-overexpressed transgenic rice.
具体实施方式Detailed ways
实验材料Experimental Materials
供试植物材料为野生型水稻(Oryza sativa)日本晴(Nipponbare)、OsC3H38过表达转基因水稻和OsC3H38RNAi转基因水稻。The tested plant materials were wild-type rice (Oryza sativa) Nipponbare, OsC3H38 overexpression transgenic rice and OsC3H38 RNAi transgenic rice.
供试菌种:大肠杆菌DH5α菌种、农杆菌EHA105菌种、酵母菌AH109。Test strains: Escherichia coli DH5α strain, Agrobacterium EHA105 strain, yeast AH109.
实施例1分析OsC3H38基因在不同处理条件下的表达模式Example 1 Analysis of the expression pattern of OsC3H38 gene under different treatment conditions
对野生型水稻进行175mM NaCl和5μM ABA处理,分别于处理0h、1h、6h、9h、12h和24h时取叶片为材料,用Trizol分别提取不同材料的总RNA,反转录后进行定量PCR分析,检测OsC3H38的转录水平。用于检测OsC3H38基因的引物对序列如下:Wild-type rice was treated with 175mM NaCl and 5μM ABA, and leaves were taken as materials at 0h, 1h, 6h, 9h, 12h, and 24h, respectively, and the total RNA of different materials was extracted with Trizol. After reverse transcription, quantitative PCR analysis was performed. , to detect the transcription level of OsC3H38. The primer pair sequences used to detect the OsC3H38 gene are as follows:
正向引物为:5′-GGTTCCACTGTTCAATCTTCGC-3′;The forward primer is: 5'-GGTTCCACTGTTCAATCTTCGC-3';
反向引物为:5′-TATGATGATGGAGGAAGGCGC-3′。The reverse primer was: 5'-TATGATGATGGAGGAAGGCGC-3'.
用于检测内参基因(actin)的表达量的引物对为:The primer pairs used to detect the expression level of the internal reference gene (actin) are:
正向引物为:5′-TGGCATCTCTCAGCACATTCC-3′;The forward primer is: 5'-TGGCATCTCTCAGCACATTCC-3';
反向引物为:5′-TGCACAATGGATGGGTCAGA-3′。The reverse primer is: 5'-TGCACAATGGATGGGTCAGA-3'.
如图1所示,OsC3H38基因在175mM NaCl处理1h时开始表达上调,至胁迫24h时,上调倍数达到5.8倍;而在5μM ABA处理后先下调表达,在处理6h时表达量开始上调表达,在胁迫12h时表达量是对照的8倍左右。因此,OsC3H38的表达受高盐和ABA诱导,是一个响应盐胁迫的耐逆相关基因。As shown in Figure 1, the expression of OsC3H38 gene started to be up-regulated after 1 h of 175 mM NaCl treatment, and the up-regulated fold reached 5.8 times when the stress was 24 h; while the expression of OsC3H38 was down-regulated first after 5 μM ABA treatment, and the expression began to be up-regulated after 6 h of treatment. Under stress for 12h, the expression level was about 8 times that of the control. Therefore, the expression of OsC3H38 is induced by high salt and ABA, and it is a stress tolerance-related gene in response to salt stress.
实施例2 OsC3H38基因的克隆及其编码蛋白的转录因子特征分析Example 2 Cloning of the OsC3H38 gene and analysis of the transcription factor characteristics of the encoded protein
1.克隆OsC3H38基因1. Cloning the OsC3H38 gene
以实施例1中的高盐胁迫12h的叶片材料的cDNA为模板,设计PCR引物序列,正向引物为:5′-AGCAAAGCGGGGATTCAG-3′,反向引物为:5′-GAACCGGCCATCCATAGAG-3′,进行RT-PCR扩增,对扩增产物进行电泳,回收、加A,与T-载体连接后转化大肠杆菌DH-5α,筛选阳性克隆后进行测序,获得测序正确的克隆命名为pT-OsC3H38,完成OsC3H38基因的克隆。Using the cDNA of the leaf material under high-salt stress for 12 h in Example 1 as a template, the PCR primer sequences were designed, the forward primer was: 5'-AGCAAAGCGGGGATTCAG-3', and the reverse primer was: 5'-GAACCGGCCATCCATAGAG-3'. RT-PCR amplification, electrophoresis of the amplified product, recovery, addition of A, ligation with T-vector, transformation of E. coli DH-5α, screening of positive clones and sequencing, the clone with correct sequencing was obtained and named pT-OsC3H38, completed Cloning of the OsC3H38 gene.
2.OsC3H38蛋白定位于细胞核里2. OsC3H38 protein is localized in the nucleus
构建OsC3H38与GFP融合表达载体p1300-OsC3H38::GFP,将该载体与对照p1300-GFP分别转化农杆菌,分别获得阳性菌后进行摇培,集菌后分别注射烟草表皮细胞,注射后培养40-48h内取叶片进行confocal观察。The OsC3H38 and GFP fusion expression vector p1300-OsC3H38::GFP was constructed, and the vector and the control p1300-GFP were transformed into Agrobacterium, respectively, and the positive bacteria were obtained and then shaken and cultured. The leaves were taken for confocal observation within 48h.
如图2所示,转化p1300-GFP的烟草表皮细胞的细胞核和细胞膜中都有绿色荧光,而转化p1300-OsC3H38::GFP的烟草中只在细胞核中发现绿色荧光,证明OsC3H38蛋白定位在细胞核中。As shown in Figure 2, green fluorescence was found in the nucleus and cell membrane of tobacco epidermal cells transformed with p1300-GFP, while only green fluorescence was found in the nucleus of tobacco transformed with p1300-OsC3H38::GFP, indicating that OsC3H38 protein was localized in the nucleus .
3.OsC3H38蛋白具有转录激活活性3. OsC3H38 protein has transcriptional activation activity
构建pGBKT7-OsC3H38质粒,转化酵母菌AH109,获得转化成功的酵母菌后,分别挑取5-6个克隆,分别用超纯水悬浮,并梯度稀释至10倍、100倍,分别取不同的悬浮液各2μL点在SD/Trp-/His-培养基上,于28℃培养箱静置培养3d后观察结果。以pGBKT7空载体的转化结果做为对照。Construct pGBKT7-OsC3H38 plasmid and transform yeast AH109. After obtaining successfully transformed yeast, pick 5-6 clones respectively, suspend them in ultrapure water, and dilute them to 10 times and 100 times in a gradient manner. 2 μL of each solution was placed on SD/Trp-/His-medium, and the results were observed after 3 days of static culture in a 28°C incubator. The transformation results of pGBKT7 empty vector were used as control.
如图3中A所示,pGBKT7和pGBKT7-OsC3H38转化的酵母菌均能在SD/Trp-培养基上正常生长;在SD/Trp-/His-培养基上pGBKT7的转化子不能生长,而pGBKT7-OsC3H38的转化子则能正常生长。上述结果说明OsC3H38蛋白能够启动报告基因His的表达。As shown in A in Figure 3, yeast transformed with pGBKT7 and pGBKT7-OsC3H38 could grow normally on SD/Trp-medium; transformants of pGBKT7 could not grow on SD/Trp-/His-medium, while pGBKT7 The transformants of -OsC3H38 could grow normally. The above results indicate that the OsC3H38 protein can initiate the expression of the reporter gene His.
分别将图3中A的一缺培养基上生长2d的包含质粒pGBKT7或pGBKT7-OsC3H38的酵母菌液点于干净滤纸上,经液氮冻融裂解后进行X-gal显色反应。The yeast solution containing the plasmid pGBKT7 or pGBKT7-OsC3H38 grown on the one-deficient medium of Figure 3 A for 2 days was spotted on clean filter paper, and then subjected to X-gal color reaction after freeze-thaw lysis with liquid nitrogen.
如图3中B所示,含pGBKT7的酵母菌没有被染蓝,而含pGBKT7-OsC3H38的酵母菌被染蓝了,表明OsC3H38蛋白成功激活LacZ报告基因的表达。As shown in B in Figure 3, the yeast containing pGBKT7 was not stained blue, while the yeast containing pGBKT7-OsC3H38 was stained blue, indicating that the OsC3H38 protein successfully activated the expression of the LacZ reporter gene.
上述结果表明OsC3H38蛋白在酵母中具有转录激活活性。The above results indicate that OsC3H38 protein has transcriptional activation activity in yeast.
因此,本发明的OsC3H38蛋白是定位在细胞核里的、具有转录激活活性的转录因子。Therefore, the OsC3H38 protein of the present invention is a transcription factor localized in the nucleus and has transcriptional activation activity.
实施例3鉴定OsC3H38的耐盐功能Example 3 Identification of the salt-tolerant function of OsC3H38
构建pMDC32-OsC3H38过表达载体和pTCK303RNAi载体,分别转化农杆菌EHA105;得到鉴定正确的转化子后,分别转化日本晴水稻,获得转基因水稻植株。分别对获得的不同转基因株系水稻进行DNA和RNA水平鉴定,鉴定结果如图4所示。鉴定后培养3-4代,获得转基因纯合株系用于以下实验。The pMDC32-OsC3H38 overexpression vector and pTCK303RNAi vector were constructed and transformed into Agrobacterium EHA105 respectively; after obtaining the correctly identified transformants, they were transformed into Nipponbare rice respectively to obtain transgenic rice plants. The DNA and RNA levels of the obtained different transgenic rice lines were identified respectively, and the identification results are shown in Figure 4. After identification, the cells were cultured for 3-4 generations, and transgenic homozygous lines were obtained for the following experiments.
1.OsC3H38过表达转基因水稻幼苗的耐逆性检测1. Detection of stress tolerance in OsC3H38 overexpressing transgenic rice seedlings
将萌发后正常生长3d的OsC3H38过表达转基因水稻株系和野生型日本晴水稻幼苗转移到含175mM NaCl的1/2MS培养基上进行处理,7d后对水稻地上部分的长度进行测量并统计数据。The OsC3H38 overexpressing transgenic rice lines and wild-type Nipponbare rice seedlings that had grown normally for 3 days after germination were transferred to 1/2MS medium containing 175 mM NaCl for treatment, and the length of the aerial part of the rice was measured and counted 7 days later.
如图5、6所示,1/2MS培养基上的对照组中3个OsC3H38过表达转基因株系、3个OsC3H38RNAi转基因株系与野生型日本晴的地上部分的长度没有显著差异;175mM NaCl处理后,3个OsC3H38过表达转基因株系的株高均高于野生型日本晴的,而3个OsC3H38RNAi转基因株系的株高均低于野生型日本晴的。因此,OsC3H38基因的上调表达增强了转基因水稻幼苗对盐胁迫的耐受性。As shown in Figures 5 and 6, the lengths of the aerial parts of the three OsC3H38 overexpression transgenic lines and the three OsC3H38 RNAi transgenic lines in the control group on 1/2 MS medium were not significantly different from those of the wild-type Nipponbare; after 175 mM NaCl treatment , the plant heights of the three OsC3H38 overexpression transgenic lines were higher than those of the wild-type Nipponbare, while the plant heights of the three OsC3H38RNAi transgenic lines were all lower than those of the wild-type Nipponbare. Therefore, the up-regulated expression of the OsC3H38 gene enhanced the tolerance of transgenic rice seedlings to salt stress.
2.OsC3H38过表达转基因水稻分蘖期耐盐性检测2. Detection of salt tolerance in tillering stage of OsC3H38 overexpressed transgenic rice
将不同株系的水稻种子萌发7d后,选取长势一致的幼苗转移到花盆中,移栽时将转基因株系和野生型对照的幼苗分别种在同一个花盆的两侧。在温室正常培养30d后改用175mM NaCl浇灌进行盐处理,盐处理18d复水6d后,观察表型。After 7 days of germination of rice seeds from different lines, seedlings with the same growth vigor were selected and transferred to flowerpots. When transplanting, seedlings of transgenic lines and wild-type control were planted on both sides of the same flowerpot. After normal cultivation in the greenhouse for 30 d, the mice were irrigated with 175 mM NaCl for salt treatment, and the phenotypes were observed after 18 d of salt treatment and rehydration for 6 d.
如图7所示,日本晴水稻茎部和叶片枯萎、变黄,OsC3H38过表达转基因水稻幼苗的叶片更绿、更挺拔;盐处理22d复水10d后相对于野生型幼苗,OsC3H38RNAi转基因水稻幼苗的叶片更加萎蔫。因此,OsC3H38基因上调表达提高了转基因水稻在分蘖期的耐盐性。As shown in Figure 7, the stems and leaves of Nipponbare rice withered and turned yellow, and the leaves of OsC3H38 overexpressed transgenic rice seedlings were greener and more upright; after salt treatment for 22 d and rehydration for 10 d, the leaves of OsC3H38RNAi transgenic rice seedlings were compared with wild-type seedlings. more wilted. Therefore, the up-regulated expression of OsC3H38 improved the salt tolerance of transgenic rice at tillering stage.
因此,OsC3H38基因上调表达提高了转基因水稻的耐盐性,是水稻耐盐的正调节子。Therefore, the up-regulated expression of OsC3H38 improves the salt tolerance of transgenic rice and is a positive regulator of salt tolerance in rice.
实施例4 OsC3H38的耐逆机理Example 4 Reversal resistance mechanism of OsC3H38
1.OsC3H38过表达改善细胞的耐逆相关生理指标1. OsC3H38 overexpression improves the physiological indicators of stress tolerance in cells
以盐胁迫处理3d的不同水稻株系为材料,进行叶绿素含量、脯氨酸含量以及丙二醛含量等耐逆相关生理指标的检测。The physiological indicators related to stress tolerance such as chlorophyll content, proline content and malondialdehyde content were detected by using different rice lines treated with salt stress for 3 days as materials.
如图8所示,盐处理条件下,过表达转基因水稻株系的叶绿素含量、脯氨酸含量和全闭气孔的比率均高于野生型日本晴的相应指标,丙二醛的含量却低于日本晴的;而OsC3H38RNAi转基因水稻的叶绿素含量、脯氨酸含量和全闭气孔的比率均低于野生型水稻的相应指标,丙二醛含量却比野生型水稻的高。As shown in Figure 8, under the condition of salt treatment, the chlorophyll content, proline content and the ratio of fully closed stomata in the overexpressed transgenic rice lines were all higher than those of the wild-type Nipponbare, but the MDA content was lower than that of Nipponbare However, the chlorophyll content, proline content and the ratio of fully closed stomata in OsC3H38RNAi transgenic rice were lower than those of wild-type rice, but the content of malondialdehyde was higher than that of wild-type rice.
因此,盐胁迫后的生理指标测定的数据与表型结果一致,OsC3H38上调表达导致了叶片细胞在生理水平发生综合改变,从而提高了水稻的耐盐能力。Therefore, the measured data of physiological indicators after salt stress are consistent with the phenotypic results. The up-regulated expression of OsC3H38 leads to comprehensive changes in leaf cells at the physiological level, thereby improving the salt tolerance of rice.
2.OsC3H38过表达提高了转基因株系中抗逆相关的标记基因的表达2. OsC3H38 overexpression increases the expression of stress resistance-related marker genes in transgenic lines
以表1中的序列为引物,利用实时定量PCR技术分别检测OsC3H38过表达株系中与非生物胁迫相关的标记基因的表达量。Using the sequences in Table 1 as primers, real-time quantitative PCR technology was used to detect the expression levels of marker genes related to abiotic stress in the OsC3H38 overexpression lines, respectively.
表1耐逆相关基因的实时定量PCR引物Table 1 Real-time quantitative PCR primers for inverse resistance-related genes

如图9所示,OsC3H38上调表达提高了OsLEA3、OsSRO1C和OsRAB21的表达量,而OsDREB1B的表达量变化不明显。OsLEA3、OsSRO1C和OsRAB21是已知的具有耐盐功能的基因,它们上调表达有利于植株耐盐。因此,OsC3H38基因从分子水平上调节了细胞的耐盐能力,有利于水稻对胁迫做出快速反应。As shown in Figure 9, the up-regulated expression of OsC3H38 increased the expression levels of OsLEA3, OsSRO1C and OsRAB21, while the expression level of OsDREB1B did not change significantly. OsLEA3, OsSRO1C and OsRAB21 are known genes with salt tolerance function, and their up-regulated expression is beneficial to plant salt tolerance. Therefore, the OsC3H38 gene regulates the salt tolerance of cells at the molecular level, which is beneficial for rice to respond quickly to stress.
序列表sequence listing
<110> 河北师范大学<110> Hebei Normal University
<120> 水稻耐盐转录因子OsC3H38及其编码基因和应用<120> Rice salt tolerance transcription factor OsC3H38 and its encoding gene and application
<160> 2<160> 2
<170> SIPOSequenceListing 1.0<170> SIPOSequenceListing 1.0
<210> 1<210> 1
<211> 304<211> 304
<212> PRT<212> PRT
<213> 水稻(Oryza sativa)<213> Rice (Oryza sativa)
<400> 1<400> 1
Met Ala Glu His Leu Ala Ser Ile Phe Gly Thr Glu Lys Asp Arg ValMet Ala Glu His Leu Ala Ser Ile Phe Gly Thr Glu Lys Asp Arg Val
1 5 10 151 5 10 15
Asn Cys Pro Phe Tyr Phe Lys Ile Gly Ala Cys Arg His Gly Asp ArgAsn Cys Pro Phe Tyr Phe Lys Ile Gly Ala Cys Arg His Gly Asp Arg
20 25 30 20 25 30
Cys Ser Arg Leu His Asn Arg Pro Thr Val Ser Pro Thr Ile Val LeuCys Ser Arg Leu His Asn Arg Pro Thr Val Ser Pro Thr Ile Val Leu
35 40 45 35 40 45
Ala Asn Met Tyr Gln Arg Pro Asp Met Ile Thr Pro Gly Val Asp AlaAla Asn Met Tyr Gln Arg Pro Asp Met Ile Thr Pro Gly Val Asp Ala
50 55 60 50 55 60
Gln Gly Gln Pro Ile Asp Pro Glu Lys Met Gln Glu His Phe Glu AspGln Gly Gln Pro Ile Asp Pro Glu Lys Met Gln Glu His Phe Glu Asp
65 70 75 8065 70 75 80
Phe Tyr Glu Asp Ile Tyr Glu Glu Leu Ser Lys Phe Gly Glu Val GluPhe Tyr Glu Asp Ile Tyr Glu Glu Leu Ser Lys Phe Gly Glu Val Glu
85 90 95 85 90 95
Thr Leu Asn Val Cys Asp Asn Leu Ala Asp His Met Ile Gly Asn ValThr Leu Asn Val Cys Asp Asn Leu Ala Asp His Met Ile Gly Asn Val
100 105 110 100 105 110
Tyr Val Gln Phe Arg Glu Glu Glu Gln Ala Val Ala Ala His Asn AlaTyr Val Gln Phe Arg Glu Glu Glu Gln Ala Val Ala Ala His Asn Ala
115 120 125 115 120 125
Leu Gln Gly Arg Phe Tyr Ser Gly Arg Pro Ile Ile Val Glu Tyr SerLeu Gln Gly Arg Phe Tyr Ser Gly Arg Pro Ile Ile Val Glu Tyr Ser
130 135 140 130 135 140
Pro Val Thr Asp Phe Arg Glu Ala Thr Cys Arg Gln Phe Glu Glu AsnPro Val Thr Asp Phe Arg Glu Ala Thr Cys Arg Gln Phe Glu Glu Asn
145 150 155 160145 150 155 160
Ser Cys Asn Arg Gly Gly Tyr Cys Asn Phe Met His Val Lys Gln IleSer Cys Asn Arg Gly Gly Tyr Cys Asn Phe Met His Val Lys Gln Ile
165 170 175 165 170 175
Gly Arg Glu Leu Arg Arg Lys Leu Tyr Gly Gly Arg Ser Arg Arg SerGly Arg Glu Leu Arg Arg Lys Leu Tyr Gly Gly Arg Ser Arg Arg Ser
180 185 190 180 185 190
His Gly Arg Ser Arg Ser Pro Ser Pro Arg His Arg Arg Gly Asn ArgHis Gly Arg Ser Arg Ser Pro Ser Pro Arg His Arg Arg Gly Asn Arg
195 200 205 195 200 205
Asp Arg Asp Asp Phe Arg Arg Glu Arg Asp Gly Tyr Arg Gly Gly GlyAsp Arg Asp Asp Phe Arg Arg Glu Arg Asp Gly Tyr Arg Gly Gly Gly
210 215 220 210 215 220
Asp Gly Tyr Arg Gly Gly Gly Gly Gly Gly Gly Gly Asp Gly Tyr ArgAsp Gly Tyr Arg Gly Gly Gly Gly Gly Gly Gly Gly Asp Gly Tyr Arg
225 230 235 240225 230 235 240
Gly Gly Asp Ser Tyr Arg Gly Gly Gly Gly Gly Gly Arg Arg Gly GlyGly Gly Asp Ser Tyr Arg Gly Gly Gly Gly Gly Gly Gly Arg Arg Gly Gly
245 250 255 245 250 255
Gly Ser Arg Tyr Asp Arg Tyr Asp Asp Gly Gly Arg Arg Arg His GlyGly Ser Arg Tyr Asp Arg Tyr Asp Asp Gly Gly Arg Arg Arg His Gly
260 265 270 260 265 270
Ser Pro Pro Arg Arg Ala Arg Ser Pro Val Arg Glu Ser Ser Glu GluSer Pro Pro Arg Arg Ala Arg Ser Pro Val Arg Glu Ser Ser Glu Glu
275 280 285 275 280 285
Arg Arg Ala Lys Ile Glu Gln Trp Asn Arg Glu Arg Glu Glu Lys ProArg Arg Ala Lys Ile Glu Gln Trp Asn Arg Glu Arg Glu Glu Lys Pro
290 295 300 290 295 300
<210> 2<210> 2
<211> 915<211> 915
<212> DNA<212> DNA
<213> 水稻(Oryza sativa)<213> Rice (Oryza sativa)
<400> 2<400> 2
atggcggagc atttggcttc aatatttggg actgagaagg atagggttaa ctgccccttt 60atggcggagc atttggcttc aatatttggg actgagaagg atagggttaa ctgccccttt 60
tacttcaaga ttggggcttg ccgtcatggg gatcgttgct cccgtctgca caacaggcca 120tacttcaaga ttggggcttg ccgtcatggg gatcgttgct cccgtctgca caacaggcca 120
actgtgtcgc cgacaattgt gcttgctaac atgtatcagc gccctgatat gatcacacca 180actgtgtcgc cgacaattgt gcttgctaac atgtatcagc gccctgatat gatcacacca 180
ggagtcgatg ctcaaggcca acctattgat cctgaaaaga tgcaggagca ctttgaggat 240ggagtcgatg ctcaaggcca acctattgat cctgaaaaga tgcaggagca ctttgaggat 240
ttctatgagg atatctatga ggagctgagc aagtttggtg aggttgagac cctcaatgtc 300ttctatgagg atatctatga ggagctgagc aagtttggtg aggttgagac cctcaatgtc 300
tgtgataacc ttgctgacca catgataggc aatgtgtatg tccagttcag ggaggaagaa 360tgtgataacc ttgctgacca catgataggc aatgtgtatg tccagttcag ggaggaagaa 360
caggcagtcg ctgcgcataa tgcccttcaa ggtcgcttct actcagggcg cccaataatt 420caggcagtcg ctgcgcataa tgcccttcaa ggtcgcttct actcagggcg cccaataatt 420
gttgagtatt cccctgtgac tgatttccgt gaggcgacct gtaggcagtt tgaggagaac 480gttgagtatt cccctgtgac tgatttccgt gaggcgacct gtaggcagtt tgaggagaac 480
agctgcaacc gtgggggcta ctgtaatttt atgcatgtaa agcagattgg cagggagctc 540agctgcaacc gtgggggcta ctgtaatttt atgcatgtaa agcagattgg cagggagctc 540
aggaggaagc tttatggggg gcgatccagg aggagccatg ggagaagccg cagcccaagc 600aggaggaagc tttatggggg gcgatccagg aggagccatg ggagaagccg cagcccaagc 600
ccacgccaca ggaggggcaa tcgtgaccgt gatgatttcc gccgtgaacg tgatggttat 660ccacgccaca ggaggggcaa tcgtgaccgt gatgatttcc gccgtgaacg tgatggttat 660
cgtggtggtg gtgatggtta ccgtggcggc ggcggtggcg gtggtggtga cggttaccgc 720cgtggtggtg gtgatggtta ccgtggcggc ggcggtggcg gtggtggtga cggttaccgc 720
ggcggtgata gttaccgtgg tggtggtggt ggtggacgca ggggtggggg cagcaggtat 780ggcggtgata gttaccgtgg tggtggtggt ggtggacgca ggggtggggg cagcaggtat 780
gacaggtatg atgatggagg aaggcgcagg catggcagcc ccccaaggcg tgcaaggagc 840gacaggtatg atgatggagg aaggcgcagg catggcagcc ccccaaggcg tgcaaggagc 840
ccagtcaggg aaagcagcga ggaacgcagg gcgaagattg aacagtggaa ccgtgaacgg 900ccagtcaggg aaagcagcga ggaacgcagg gcgaagattg aacagtggaa ccgtgaacgg 900
gaggaaaagc cttga 915gaggaaaagc cttga 915
Claims (1)
Translated fromChineseApplications Claiming Priority (2)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN2018115058925 | 2018-12-10 | ||
| CN201811505892 | 2018-12-10 |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN110028564A CN110028564A (en) | 2019-07-19 |
| CN110028564Btrue CN110028564B (en) | 2022-04-08 |
Family
ID=67236397
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201910219494.5AExpired - Fee RelatedCN110028564B (en) | 2018-12-10 | 2019-03-20 | Rice salt tolerance transcription factor OsC3H38 and its encoding gene and application |
Country Status (1)
| Country | Link |
|---|---|
| CN (1) | CN110028564B (en) |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2659414A1 (en)* | 2006-08-02 | 2008-02-07 | Cropdesign N.V. | Use of synovial sarcoma translocation (syt) polypeptide for increasing plant yield under abiotic stress |
| CN104480120A (en)* | 2014-12-11 | 2015-04-01 | 中国农业科学院生物技术研究所 | Plant salt-tolerant related gene PpSIG2 as well as encoding protein and application thereof |
- 2019
- 2019-03-20CNCN201910219494.5Apatent/CN110028564B/ennot_activeExpired - Fee Related
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CA2659414A1 (en)* | 2006-08-02 | 2008-02-07 | Cropdesign N.V. | Use of synovial sarcoma translocation (syt) polypeptide for increasing plant yield under abiotic stress |
| CN104480120A (en)* | 2014-12-11 | 2015-04-01 | 中国农业科学院生物技术研究所 | Plant salt-tolerant related gene PpSIG2 as well as encoding protein and application thereof |
Non-Patent Citations (3)
| Title |
|---|
| PREDICTED: Oryza sativa Japonica Group splicing factor U2af small subunit B (LOC4339621), transcript variant X1, mRNA;XM_015784235;《GenBank Database》;20180807;第1-2页* |
| splicing factor U2af small subunit B [Oryza sativa Japonica Group];XP_015639721;《GenBank Database》;20180807;第1-2页* |
| 植物耐盐性及耐盐相关基因的研究进展;邴雷;《河北师范大学学报(自然科学版)》;20080320;第32卷(第2期);第243-248页* |
Also Published As
| Publication number | Publication date |
|---|---|
| CN110028564A (en) | 2019-07-19 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| Qiu et al. | Over-expression of the stress-induced OsWRKY45 enhances disease resistance and drought tolerance in Arabidopsis | |
| Liu et al. | TCP10, a TCP transcription factor in moso bamboo (Phyllostachys edulis), confers drought tolerance to transgenic plants | |
| Zhang et al. | Overexpression of a NF-YB3 transcription factor from Picea wilsonii confers tolerance to salinity and drought stress in transformed Arabidopsis thaliana | |
| CN102234320B (en) | Plant stress-tolerant associated protein TaDREB4B and encoding gene and application thereof | |
| Lian et al. | MdDREB2A in apple is involved in the regulation of multiple abiotic stress responses | |
| CN105861517B (en) | A kind of Radix Notoginseng antibacterial peptide gene PnSN1 and its application | |
| Wang et al. | Overexpression of ERF96, a small ethylene response factor gene, enhances salt tolerance in Arabidopsis | |
| CN110872598B (en) | Cotton drought-resistant related gene GhDT1 and application thereof | |
| WO2013033571A1 (en) | Plants with enhanced tolerance to multiple abiotic stresses | |
| Tao et al. | SlDREB2 gene specifically recognizing to the universal DRE elements is a transcriptional activator improving drought tolerance in tomato | |
| CN107164391A (en) | A kind of strawberry floral genes FvbHLH78 and its application | |
| CN104892741B (en) | Plant stress tolerance correlative protein GmNF YA17 and its encoding gene and application | |
| CN111454972A (en) | The cold resistance gene PtrBADH of Citrus aurantium and its application in genetic improvement of plant cold resistance | |
| Bu et al. | The transcription factor TabZIP156 acts as a positive regulator in response to drought tolerance in Arabidopsis and wheat (Triticum aestivum L.) | |
| CN103319583B (en) | Plant stress tolerance-associated protein TaNF-YB 1, coding genes thereof and applications | |
| CN109485705B (en) | Rice drought tolerance-related transcription factor OsTLP6 and its encoding gene and application | |
| CN104220596A (en) | Plant body showing improved resistance against environmental stress and method for producing same | |
| CN102234322B (en) | Plant stress tolerance related protein GmNF-YA1 and its coding gene and application | |
| CN110922461B (en) | Rice drought tolerance-related transcription factor OsAE10 and its encoding gene and application | |
| CN116497038B (en) | A cold-resistant gene MfJAZ1 of yellow alfalfa and its application | |
| CN104892737B (en) | Plant stress tolerance correlative protein GmNF-YA15 and its encoding gene and application | |
| CN108409846B (en) | A Soybean Salt Tolerance-Related MYB Transcription Factor and Its Encoding Gene and Application | |
| CN110028564B (en) | Rice salt tolerance transcription factor OsC3H38 and its encoding gene and application | |
| CN113666993B (en) | Alfalfa MsSPL12 protein and related biological materials and their application in improving plant stress resistance | |
| CN106399327A (en) | Western balsam poplar PtrZFP103 gene as well as coding protein and application thereof |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| GR01 | Patent grant | ||
| GR01 | Patent grant | ||
| CF01 | Termination of patent right due to non-payment of annual fee | Granted publication date:20220408 | |
| CF01 | Termination of patent right due to non-payment of annual fee |