CN108441520B - Conditional gene knockout method constructed by CRISPR/Cas9 system - Google Patents
Conditional gene knockout method constructed by CRISPR/Cas9 systemDownload PDFInfo
- Publication number
- CN108441520B CN108441520BCN201810300747.7ACN201810300747ACN108441520BCN 108441520 BCN108441520 BCN 108441520BCN 201810300747 ACN201810300747 ACN 201810300747ACN 108441520 BCN108441520 BCN 108441520B
- Authority
- CN
- China
- Prior art keywords
- oxp
- frt
- gene
- antibiotic
- target
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 108091033409CRISPRProteins0.000titleclaimsabstractdescription43
- 238000000034methodMethods0.000titleclaimsabstractdescription30
- 238000010354CRISPR gene editingMethods0.000titleclaimsabstractdescription13
- 238000003209gene knockoutMethods0.000titleclaimsdescription17
- 108090000623proteins and genesProteins0.000claimsabstractdescription70
- 239000013612plasmidSubstances0.000claimsabstractdescription59
- 230000008685targetingEffects0.000claimsabstractdescription52
- 230000006801homologous recombinationEffects0.000claimsabstractdescription28
- 238000002744homologous recombinationMethods0.000claimsabstractdescription28
- 108020005004Guide RNAProteins0.000claimsabstractdescription26
- 230000001404mediated effectEffects0.000claimsabstractdescription24
- 239000012634fragmentSubstances0.000claimsabstractdescription14
- 239000013613expression plasmidSubstances0.000claimsabstractdescription10
- 238000012216screeningMethods0.000claimsabstractdescription8
- 238000001890transfectionMethods0.000claimsabstractdescription8
- 239000003242anti bacterial agentSubstances0.000claimsabstractdescription6
- 229940088710antibiotic agentDrugs0.000claimsabstractdescription6
- 230000001939inductive effectEffects0.000claimsabstractdescription6
- 210000004027cellAnatomy0.000claimsdescription59
- 238000003780insertionMethods0.000claimsdescription13
- 230000037431insertionEffects0.000claimsdescription13
- 108010014303DNA-directed DNA polymeraseProteins0.000claimsdescription9
- 102000016928DNA-directed DNA polymeraseHuman genes0.000claimsdescription9
- AOJJSUZBOXZQNB-TZSSRYMLSA-NDoxorubicinChemical compoundO([C@H]1C[C@@](O)(CC=2C(O)=C3C(=O)C=4C=CC=C(C=4C(=O)C3=C(O)C=21)OC)C(=O)CO)[C@H]1C[C@H](N)[C@H](O)[C@H](C)O1AOJJSUZBOXZQNB-TZSSRYMLSA-N0.000claimsdescription8
- RXWNCPJZOCPEPQ-NVWDDTSBSA-NpuromycinChemical compoundC1=CC(OC)=CC=C1C[C@H](N)C(=O)N[C@H]1[C@@H](O)[C@H](N2C3=NC=NC(=C3N=C2)N(C)C)O[C@@H]1CORXWNCPJZOCPEPQ-NVWDDTSBSA-N0.000claimsdescription8
- 230000003115biocidal effectEffects0.000claimsdescription7
- 229960004927neomycinDrugs0.000claimsdescription7
- DODQJNMQWMSYGS-QPLCGJKRSA-N4-[(z)-1-[4-[2-(dimethylamino)ethoxy]phenyl]-1-phenylbut-1-en-2-yl]phenolChemical compoundC=1C=C(O)C=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=CC=C1DODQJNMQWMSYGS-QPLCGJKRSA-N0.000claimsdescription6
- 108700028369AllelesProteins0.000claimsdescription6
- 229930193140NeomycinNatural products0.000claimsdescription6
- YQYJSBFKSSDGFO-UHFFFAOYSA-NEpihygromycinNatural productsOC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1OYQYJSBFKSSDGFO-UHFFFAOYSA-N0.000claimsdescription4
- 229950010131puromycinDrugs0.000claimsdescription4
- 230000033228biological regulationEffects0.000claimsdescription3
- 229960004679doxorubicinDrugs0.000claimsdescription3
- 101000865057Thermococcus litoralis DNA polymeraseProteins0.000claimsdescription2
- 230000003321amplificationEffects0.000claimsdescription2
- 210000004962mammalian cellAnatomy0.000claimsdescription2
- 108091027963non-coding RNAProteins0.000claimsdescription2
- 102000042567non-coding RNAHuman genes0.000claimsdescription2
- 238000003199nucleic acid amplification methodMethods0.000claimsdescription2
- 229940009456adriamycinDrugs0.000claims1
- 238000001976enzyme digestionMethods0.000claims1
- 229920002477rna polymerPolymers0.000claims1
- 125000006850spacer groupChemical group0.000claims1
- 230000009466transformationEffects0.000claims1
- 108700026220vif GenesProteins0.000claims1
- TXUZVZSFRXZGTL-QPLCGJKRSA-NafimoxifeneChemical compoundC=1C=CC=CC=1C(/CC)=C(C=1C=CC(OCCN(C)C)=CC=1)/C1=CC=C(O)C=C1TXUZVZSFRXZGTL-QPLCGJKRSA-N0.000abstractdescription7
- 101150036876cre geneProteins0.000description18
- 108020004414DNAProteins0.000description15
- 101150028000EED geneProteins0.000description13
- 210000001671embryonic stem cellAnatomy0.000description11
- 238000005516engineering processMethods0.000description11
- 238000005215recombinationMethods0.000description11
- 230000006798recombinationEffects0.000description10
- 241001465754MetazoaSpecies0.000description8
- 238000011144upstream manufacturingMethods0.000description8
- 108010051219Cre recombinaseProteins0.000description7
- 101150045486SRCAP geneProteins0.000description7
- 238000000746purificationMethods0.000description7
- 210000001519tissueAnatomy0.000description7
- 239000013598vectorSubstances0.000description7
- CURLTUGMZLYLDI-UHFFFAOYSA-NCarbon dioxideChemical compoundO=C=OCURLTUGMZLYLDI-UHFFFAOYSA-N0.000description6
- 108010091086RecombinasesProteins0.000description6
- 102000018120RecombinasesHuman genes0.000description6
- 238000010276constructionMethods0.000description6
- 238000013461designMethods0.000description6
- 238000003205genotyping methodMethods0.000description6
- 102000004190EnzymesHuman genes0.000description5
- 108090000790EnzymesProteins0.000description5
- 108091027544Subgenomic mRNAProteins0.000description5
- 230000006870functionEffects0.000description5
- 238000010362genome editingMethods0.000description5
- 210000000056organAnatomy0.000description5
- 108091008146restriction endonucleasesProteins0.000description5
- 230000014509gene expressionEffects0.000description4
- 230000009261transgenic effectEffects0.000description4
- 101000704158Homo sapiens Helicase SRCAPProteins0.000description3
- 230000008901benefitEffects0.000description3
- 229910002092carbon dioxideInorganic materials0.000description3
- 239000001569carbon dioxideSubstances0.000description3
- 238000012217deletionMethods0.000description3
- 230000037430deletionEffects0.000description3
- 230000034431double-strand break repair via homologous recombinationEffects0.000description3
- 229960003722doxycyclineDrugs0.000description3
- XQTWDDCIUJNLTR-CVHRZJFOSA-Ndoxycycline monohydrateChemical compoundO.O=C1C2=C(O)C=CC=C2[C@H](C)[C@@H]2C1=C(O)[C@]1(O)C(=O)C(C(N)=O)=C(O)[C@@H](N(C)C)[C@@H]1[C@H]2OXQTWDDCIUJNLTR-CVHRZJFOSA-N0.000description3
- 230000008439repair processEffects0.000description3
- 102100025752CASP8 and FADD-like apoptosis regulatorHuman genes0.000description2
- 108700024394ExonProteins0.000description2
- 102100031880Helicase SRCAPHuman genes0.000description2
- 101000914211Homo sapiens CASP8 and FADD-like apoptosis regulatorProteins0.000description2
- 101710163270NucleaseProteins0.000description2
- 108091028043Nucleic acid sequenceProteins0.000description2
- 238000003776cleavage reactionMethods0.000description2
- 238000010367cloningMethods0.000description2
- 230000006872improvementEffects0.000description2
- 230000010354integrationEffects0.000description2
- 239000002773nucleotideSubstances0.000description2
- 125000003729nucleotide groupChemical group0.000description2
- 230000037361pathwayEffects0.000description2
- 230000004962physiological conditionEffects0.000description2
- 230000008569processEffects0.000description2
- 102000004169proteins and genesHuman genes0.000description2
- 239000012264purified productSubstances0.000description2
- 230000008263repair mechanismEffects0.000description2
- 238000011160researchMethods0.000description2
- 230000007017scissionEffects0.000description2
- 238000012163sequencing techniqueMethods0.000description2
- 208000031873Animal Disease ModelsDiseases0.000description1
- 241000894006BacteriaSpecies0.000description1
- 238000010356CRISPR-Cas9 genome editingMethods0.000description1
- 241000282693CercopithecidaeSpecies0.000description1
- 208000031639Chromosome DeletionDiseases0.000description1
- 102000053602DNAHuman genes0.000description1
- 230000007035DNA breakageEffects0.000description1
- 230000005778DNA damageEffects0.000description1
- 231100000277DNA damageToxicity0.000description1
- 206010059866Drug resistanceDiseases0.000description1
- 102100031780EndonucleaseHuman genes0.000description1
- 108010042407EndonucleasesProteins0.000description1
- 241000588724Escherichia coliSpecies0.000description1
- 241000702191Escherichia virus P1Species0.000description1
- 101000934888Homo sapiens Succinate dehydrogenase cytochrome b560 subunit, mitochondrialProteins0.000description1
- 241000124008MammaliaSpecies0.000description1
- 108091092724Noncoding DNAProteins0.000description1
- 241000700159RattusSpecies0.000description1
- 108010052160Site-specific recombinaseProteins0.000description1
- 102100025393Succinate dehydrogenase cytochrome b560 subunit, mitochondrialHuman genes0.000description1
- 241000282887SuidaeSpecies0.000description1
- 230000002159abnormal effectEffects0.000description1
- 150000001413amino acidsChemical class0.000description1
- 238000011558animal model by diseaseMethods0.000description1
- 230000003197catalytic effectEffects0.000description1
- 238000004113cell cultureMethods0.000description1
- 230000030833cell deathEffects0.000description1
- 230000004663cell proliferationEffects0.000description1
- 230000001413cellular effectEffects0.000description1
- 238000006243chemical reactionMethods0.000description1
- 230000034994deathEffects0.000description1
- 238000011161developmentMethods0.000description1
- 201000010099diseaseDiseases0.000description1
- 208000037265diseases, disorders, signs and symptomsDiseases0.000description1
- 229940079593drugDrugs0.000description1
- 239000003814drugSubstances0.000description1
- 210000002257embryonic structureAnatomy0.000description1
- 239000003623enhancerSubstances0.000description1
- 238000002474experimental methodMethods0.000description1
- 239000013604expression vectorSubstances0.000description1
- 231100000221frame shift mutation inductionToxicity0.000description1
- 230000037433frameshiftEffects0.000description1
- 238000012239gene modificationMethods0.000description1
- 230000005017genetic modificationEffects0.000description1
- 235000013617genetically modified foodNutrition0.000description1
- PCHJSUWPFVWCPO-UHFFFAOYSA-NgoldChemical compound[Au]PCHJSUWPFVWCPO-UHFFFAOYSA-N0.000description1
- 239000010931goldSubstances0.000description1
- 229910052737goldInorganic materials0.000description1
- 238000009396hybridizationMethods0.000description1
- 230000006698inductionEffects0.000description1
- 230000001665lethal effectEffects0.000description1
- 230000007246mechanismEffects0.000description1
- 230000004048modificationEffects0.000description1
- 238000012986modificationMethods0.000description1
- 230000006780non-homologous end joiningEffects0.000description1
- 231100000683possible toxicityToxicity0.000description1
- 239000000047productSubstances0.000description1
- 238000009790rate-determining step (RDS)Methods0.000description1
- 238000011084recoveryMethods0.000description1
- 230000001105regulatory effectEffects0.000description1
- 230000001568sexual effectEffects0.000description1
- 230000002123temporal effectEffects0.000description1
Images





Classifications
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/87—Introduction of foreign genetic material using processes not otherwise provided for, e.g. co-transformation
- C12N15/90—Stable introduction of foreign DNA into chromosome
- C12N15/902—Stable introduction of foreign DNA into chromosome using homologous recombination
- C12N15/907—Stable introduction of foreign DNA into chromosome using homologous recombination in mammalian cells
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/85—Vectors or expression systems specially adapted for eukaryotic hosts for animal cells
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N9/00—Enzymes; Proenzymes; Compositions thereof; Processes for preparing, activating, inhibiting, separating or purifying enzymes
- C12N9/14—Hydrolases (3)
- C12N9/16—Hydrolases (3) acting on ester bonds (3.1)
- C12N9/22—Ribonucleases [RNase]; Deoxyribonucleases [DNase]
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/10—Plasmid DNA
- C12N2800/106—Plasmid DNA for vertebrates
- C12N2800/107—Plasmid DNA for vertebrates for mammalian
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2800/00—Nucleic acids vectors
- C12N2800/30—Vector systems comprising sequences for excision in presence of a recombinase, e.g. loxP or FRT
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N2810/00—Vectors comprising a targeting moiety
- C12N2810/10—Vectors comprising a non-peptidic targeting moiety
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Engineering & Computer Science (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Bioinformatics & Cheminformatics (AREA)
- Wood Science & Technology (AREA)
- Zoology (AREA)
- Biomedical Technology (AREA)
- Biotechnology (AREA)
- General Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Microbiology (AREA)
- Biochemistry (AREA)
- General Health & Medical Sciences (AREA)
- Physics & Mathematics (AREA)
- Biophysics (AREA)
- Plant Pathology (AREA)
- Medicinal Chemistry (AREA)
- Cell Biology (AREA)
- Mycology (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Description
Translated fromChinese技术领域technical field
本发明涉及基因敲除技术领域,尤其涉及一种利用CRISPR/Cas9和Cre-LoxP系统构建的基因条件性敲除方法。The invention relates to the technical field of gene knockout, in particular to a gene conditional knockout method constructed by using CRISPR/Cas9 and Cre-LoxP systems.
背景技术Background technique
基因敲除技术是转基因动物构建中不可替代的组成部分。基因的表达具有时间和空间特异性,对体内关键基因进行敲除和替换等方式的基因编辑,往往导致动物胚胎死亡,阻碍基因的功能研究。条件性敲除方法的出现解决了上述问题,可以使目标基因的表达或缺失发生在动物发育的某一特定阶段或某一特定组织器官。条件性基因敲除技术作为新一代基因敲除技术,能够实现基因在空间和时间上的特异性表达,相对于组成型基因敲除有着明显的优势和广泛的应用前景。Gene knockout technology is an irreplaceable component in the construction of transgenic animals. Gene expression is temporal and spatial specific. Gene editing in the form of knockout and replacement of key genes in the body often leads to the death of animal embryos and hinders the functional study of genes. The emergence of conditional knockout methods solves the above problems, which can make the expression or deletion of target genes occur in a specific stage of animal development or a specific tissue organ. Conditional gene knockout technology, as a new generation gene knockout technology, can realize the specific expression of genes in space and time, and has obvious advantages and broad application prospects compared with constitutive gene knockout.
传统的基因条件性敲除方法利用在目的敲除序列的两端插入位点特异性的重组酶识别位点,在重组酶的诱导下,通过两侧序列的缺失或倒置获得无效等位基因。因此,重组酶介导的条件性敲除成为研究特异性细胞类型和不同发育阶段中的基因功能极有价值的工具。但是这种条件性敲除技术在构建载体时需要消耗大量的时间和精力,不仅要利用BAC克隆和基因重组等技术,基因型鉴定还需要长片段PCR、Northern等方法,而且基因敲除效率也相对较低。The traditional gene conditional knockout method utilizes the insertion of site-specific recombinase recognition sites at both ends of the target knockout sequence, and under the induction of the recombinase, the null alleles are obtained by deletion or inversion of the sequences on both sides. Therefore, recombinase-mediated conditional knockout becomes an extremely valuable tool to study gene function in specific cell types and different developmental stages. However, this conditional knockout technology requires a lot of time and energy when constructing a vector. Not only BAC cloning and gene recombination techniques are used, but also long-fragment PCR, Northern and other methods are required for genotype identification, and the gene knockout efficiency is also relatively low.
目前主要基于Cre-LoxP系统对基因进行条件性敲除,该系统包含Cre重组酶和loxP序列。Cre重组酶是大肠杆菌噬菌体P1中cre基因编码表达的由343个氨基酸构成的蛋白质,它不仅具有催化活性,而且与限制酶相似,能够特异性识别细胞内基因或DNA上的loxP序列,并根据loxP序列的位置和序列之间的方向介导DNA序列发生特异性重组反应。Cre重组酶是一种比较稳定的蛋白质,可以在生物体不同的组织、不同的生理条件下发挥作用,因此Cre重组酶的编码基因可以置于组织、器官特异的启动子的调控之下,诱导重组酶在生物体不同的细胞、组织、器官,以及不同的发育阶段或不同的生理条件下表达,进而发挥作用。loxP是一段长34bp的DNA序列,含有两个13bp的反向重复序列和一个8bp的核心序列。任何序列的DNA,当其位于两个loxP位点之间的时候,在Cre重组酶的作用下要么被缺失(两个loxP位点的方向相同),要么方向发生倒转(两loxP位点的方向相反)。At present, conditional knockout of genes is mainly based on the Cre-LoxP system, which includes Cre recombinase and loxP sequences. Cre recombinase is a protein composed of 343 amino acids encoded and expressed by the cre gene in Escherichia coli phage P1. It not only has catalytic activity, but also is similar to restriction enzymes. It can specifically recognize loxP sequences on genes or DNA in cells, and according to the The position and orientation of loxP sequences mediate specific recombination reactions of DNA sequences. Cre recombinase is a relatively stable protein that can function in different tissues and physiological conditions of the organism. Therefore, the gene encoding Cre recombinase can be placed under the regulation of tissue and organ-specific promoters to induce Recombinases are expressed in different cells, tissues, and organs of organisms, as well as at different developmental stages or under different physiological conditions, and then play their role. loxP is a DNA sequence of 34 bp in length, containing two 13 bp inverted repeats and an 8 bp core sequence. DNA of any sequence, when it is located between two loxP sites, is either deleted by Cre recombinase (the two loxP sites are in the same direction) or the direction is reversed (the two loxP sites are in the same direction) on the contrary).
基于Cre/loxP系统的基因条件性敲除技术需要两步:(1)在细胞的基因组中引入loxP序列,靶向载体的构建和对同源重组细胞的筛选来实现。(2)通过Cre介导的重组来实现靶基因编辑。Cre/loxP系统既可以在细胞水平上用Cre重组酶识别loxP位点将靶基因切除,同时可以在个体水平上将含有loxP的转基因动物与Cre转基因动物杂交,得到目的基因条件性敲除得动物,或者将Cre基因置于可诱导的启动子控制下,通过诱导表达Cre重组酶而将loxP位点之间的基因切除(诱导性基因敲除),实现目的基因在特定时间或者特定组织中的敲除。Cre/loxP系统构建的基因条件性敲除细胞或转基因动物,避免了某些基因完全敲除所导致的胚胎致死效应,现已广泛应用于免疫学、动物疾病模型构建及疾病相关基因功能鉴定等领域。The gene conditional knockout technology based on the Cre/loxP system requires two steps: (1) The loxP sequence is introduced into the genome of the cell, the construction of the targeting vector and the screening of the homologous recombination cells are realized. (2) Target gene editing is achieved through Cre-mediated recombination. The Cre/loxP system can not only use Cre recombinase to recognize the loxP site to excise the target gene at the cellular level, but also cross the transgenic animals containing loxP with Cre transgenic animals at the individual level to obtain animals with conditional knockout of the target gene , or place the Cre gene under the control of an inducible promoter, and excise the gene between the loxP sites by inducing the expression of Cre recombinase (inducible gene knockout) to achieve the target gene at a specific time or in a specific tissue. knockout. The gene conditional knockout cells or transgenic animals constructed by the Cre/loxP system avoid the embryonic lethal effect caused by the complete knockout of some genes, and have been widely used in immunology, animal disease model construction and disease-related gene function identification, etc. field.
虽然Cre/loxP系统在理论上已十分成熟但在实际应用中仍存在不足和限制:Although the Cre/loxP system is very mature in theory, there are still shortcomings and limitations in practical applications:
(1)利用Cre/loxP技术进行基因的条件性敲除首先要构建打靶载体,扩增长度约为4-5kb的同源臂,同时还需要BAC克隆和基因重组技术。将loxP片段靶向导入细胞基因组,这一步发生同源重组的概率极低,并且需要通过长片段PCR和Northern杂交等复杂的筛选工作,才能鉴定靶向成功的细胞。因此,该步骤需要大量的时间和精力,是利用Cre/loxP系统进行基因条件性敲除的限速步骤。(1) Conditional knockout of genes using Cre/loxP technology firstly constructs a targeting vector, amplifies homology arms with a length of about 4-5kb, and also requires BAC cloning and gene recombination technology. Targeted introduction of loxP fragments into the cell genome, the probability of homologous recombination in this step is extremely low, and complex screening work such as long-range PCR and Northern hybridization is required to identify successfully targeted cells. Therefore, this step requires a lot of time and effort and is the rate-limiting step for conditional gene knockout using the Cre/loxP system.
(2)Cre酶作为一种外源性重组酶在哺乳动物中表达后具有一定的潜在毒性,会引起细胞增殖异常、DNA错配和染色体缺失等问题。(2) Cre enzyme, as an exogenous recombinase, has certain potential toxicity after being expressed in mammals, which can cause problems such as abnormal cell proliferation, DNA mismatch and chromosome deletion.
(3)Cre/loxP系统切割效率在70%左右,目的器官或组织中仍有少量敲除基因未被敲除,这些未敲除的目的基因会在特定器官或组织内微量表达。(3) The cleavage efficiency of the Cre/loxP system is about 70%, and there are still a few knockout genes that have not been knocked out in the target organs or tissues, and these target genes that are not knocked out will be slightly expressed in specific organs or tissues.
CRISPR/Cas9技术是一种新型的基因编辑技术,是动物遗传改造和基因功能研究的重要工具。CRISPR/Cas9技术主要通过核酸酶Cas9起作用,Cas9在化学合成或载体表达的sgRNA(向导RNA)引导下能够对靶基因进行切割,造成靶基因双链DNA断裂。DNA损伤可以启动细胞内的修复机制,一种是低保真性的非同源末端连接途径(NHEJ,Non-homologous endjoining),此修复机制非常容易发生错误,导致修复后发生碱基的缺失或插入,从而造成移码突变,最终达到基因敲除的目的。第二种DNA断裂修复途径为同源介导的修复(HR,homology-directed repair),这种基于同源重组的修复机制保真性高,但发生概率很低,在提供外源修复模板的情况下,靶向核酸酶对DNA的切割后可以大幅提高同源重组效率。CRISPR-Cas9系统的RNA-DNA识别机制为基因组工程研究提供了一项简便而强大的工具,该体系另一个重要的优势是Cas9蛋白可在多个不同的gRNA的引导下同时靶向多个基因组位点,CRISPR-Cas系统的高效基因组编辑功能已被应用于多种生物,包括猴子、猪、大鼠、秀丽隐杆线虫、植物及细菌。CRISPR/Cas9 technology is a new type of gene editing technology and an important tool for animal genetic modification and gene function research. CRISPR/Cas9 technology mainly works through the nuclease Cas9, which can cut the target gene under the guidance of chemically synthesized or vector-expressed sgRNA (guide RNA), resulting in double-stranded DNA breakage of the target gene. DNA damage can initiate repair mechanisms in cells, one is the low-fidelity non-homologous endjoining pathway (NHEJ), which is very error-prone, resulting in base deletions or insertions after repair , resulting in frameshift mutation, and finally achieve the purpose of gene knockout. The second DNA break repair pathway is homology-directed repair (HR, homology-directed repair). This repair mechanism based on homologous recombination has high fidelity, but the probability of occurrence is very low. In the case of providing an exogenous repair template Under the following conditions, the cleavage of DNA by targeted nucleases can greatly improve the efficiency of homologous recombination. The RNA-DNA recognition mechanism of the CRISPR-Cas9 system provides a simple and powerful tool for genome engineering research. Another important advantage of this system is that the Cas9 protein can simultaneously target multiple genomes under the guidance of multiple different gRNAs. The high-efficiency genome editing capabilities of the CRISPR-Cas system have been applied to a variety of organisms, including monkeys, pigs, rats, C. elegans, plants, and bacteria.
总之,传统的重组酶介导的基因条件性敲除,其采用自然条件下基因重组,重组效率低。传统的Cre/loxP系统将loxP位点插入到基因组中,同源臂长,发生同源重组效率低,极大影响了基因条件性敲除的效率。In conclusion, the traditional recombinase-mediated gene conditional knockout adopts gene recombination under natural conditions, and the recombination efficiency is low. The traditional Cre/loxP system inserts the loxP site into the genome, the homology arm is long, and the homologous recombination efficiency is low, which greatly affects the efficiency of conditional gene knockout.
发明内容SUMMARY OF THE INVENTION
为解决上述技术问题,本发明的目的是提供一种基于CRISPR/Cas9系统构建的基因条件性敲除方法,本发明简化并且加速了基因条件性敲除的过程,极大的提高了传统基因条件性敲除的效率和便利性。In order to solve the above technical problems, the purpose of the present invention is to provide a gene conditional knockout method based on the CRISPR/Cas9 system. The present invention simplifies and accelerates the process of gene conditional knockout, and greatly improves the traditional gene condition. Efficiency and convenience of sexual knockout.
本发明提供了一种利用CRISPR/Cas9系统构建的基因条件性敲除方法,包括以下步骤:The present invention provides a gene conditional knockout method constructed by using the CRISPR/Cas9 system, comprising the following steps:
(1)构建目的基因左侧打靶质粒和右侧打靶质粒:(1) Construct the left-side targeting plasmid and the right-side targeting plasmid of the target gene:
在敲除区域左侧和右侧分别选取LoxP-抗生素-LoxP序列待插入位点和FRT-抗生素-FRT-LoxP序列待插入位点;然后在待插入位点的上下游各选取500bp-1kb的左右同源臂,通过PCR方法扩增左右同源臂并纯化,利用限制性内切酶酶切、连接和转化等步骤,将左臂和右臂片段分别插入到LoxP-抗生素-LoxP或FRT-抗生素-FRT-LoxP打靶质粒中,得到左侧打靶质粒和右侧打靶质粒;Select the LoxP-antibiotic-LoxP sequence to be inserted and the FRT-antibiotic-FRT-LoxP sequence to be inserted on the left and right sides of the knockout region respectively; then select 500bp-1kb upstream and downstream of the to-be-inserted site The left and right homology arms are amplified and purified by PCR method, and the left and right arm fragments are inserted into LoxP-antibiotic-LoxP or FRT- In the antibiotic-FRT-LoxP targeting plasmid, the left targeting plasmid and the right targeting plasmid are obtained;
(2)通过网站http://crispr.mit.edu:8079/,设计左侧打靶质粒相对应的第一gRNA和/或右侧打靶质粒相对应的第二gRNA;(2) Through the website http://crispr.mit.edu:8079/, design the first gRNA corresponding to the targeting plasmid on the left and/or the second gRNA corresponding to the targeting plasmid on the right;
(3)将Cas9质粒、第一gRNA表达质粒和左侧打靶质粒同时转入到目的细胞中,转染48-72小时后用抗生素进行筛选,处理5-9天后,通过PCR基因型鉴定的方法,筛选出双等位基因导入LoxP-抗生素-LoxP片段的目的细胞;(3) The Cas9 plasmid, the first gRNA expression plasmid and the left-side targeting plasmid are simultaneously transferred into the target cells, screened with antibiotics 48-72 hours after transfection, and 5-9 days after treatment, the method of identifying the genotype by PCR , screen out the target cells with biallelic introduction of LoxP-antibiotic-LoxP fragment;
(4)用4-羟基他莫昔芬(4-OHT)处理双等位基因导入LoxP-抗生素-LoxP片段的目的细胞3-6天,以诱导Cre-LoxP介导的同源重组,从而使LoxP-抗生素-LoxP片段仅剩余LoxP位点,得到出LoxP双等位基因导入的目的细胞;(4) The target cells into which the biallelic LoxP-antibiotic-LoxP fragment was introduced were treated with 4-hydroxytamoxifen (4-OHT) for 3-6 days to induce Cre-LoxP-mediated homologous recombination, thereby making The LoxP-antibiotic-LoxP fragment has only the remaining LoxP site, and the target cells into which the LoxP biallelic gene is introduced are obtained;
(5)将Cas9质粒、第二gRNA表达质粒和右侧打靶质粒同时转入步骤(4)得到的LoxP双等位基因导入的目的细胞中,转染24-72小时后用抗生素进行细胞选择,选择5-9天后,筛选出插入FRT-抗生素-FRT-LoxP片段双等位基因导入的目的细胞;(5) The Cas9 plasmid, the second gRNA expression plasmid and the right-side targeting plasmid were simultaneously transferred into the target cells into which the LoxP biallelic gene obtained in step (4) was introduced, and the cells were selected with antibiotics after 24-72 hours of transfection, After 5-9 days of selection, screen out the target cells into which the FRT-antibiotic-FRT-LoxP fragment biallele was inserted;
(6)用阿霉素(Doxycycline)处理步骤(5)得到的细胞3-6天,以诱导FLP-FRT介导的同源重组,筛选出待敲除区域右侧FRT-LoxP双等位基因靶向导入的目的细胞,待敲除区域两侧被LoxP环绕,简称为Floxed细胞;(6) Treat the cells obtained in step (5) with doxorubicin (Doxycycline) for 3-6 days to induce FLP-FRT-mediated homologous recombination, and screen out the FRT-LoxP biallelic gene on the right side of the region to be knocked out The target cells that are targeted for introduction are surrounded by LoxP on both sides of the region to be knocked out, referred to as Floxed cells;
(7)用4-羟基他莫昔芬处理步骤(6)处理后的细胞3-6天,诱导Cre-LoxP介导的同源重组,以敲除目的基因。(7) Treat the cells treated in step (6) with 4-hydroxytamoxifen for 3-6 days to induce Cre-LoxP-mediated homologous recombination to knock out the target gene.
进一步地,在步骤(1)中,敲除区域为目的基因的调控区域(启动子、增强子等),非编码RNA或目的基因上的基因间隔区等DNA片段。Further, in step (1), the knockout region is the regulatory region (promoter, enhancer, etc.) of the target gene, non-coding RNA or DNA fragments such as intergenic regions on the target gene.
进一步地,在步骤(1)中,左臂利用KpnⅠ和EcoRⅠ限制性内切酶进行酶切,右臂用BamHⅠ和SacⅡ进行酶切。Further, in step (1), the left arm is digested with KpnI and EcoRI restriction enzymes, and the right arm is digested with BamHI and SacII.
进一步地,在步骤(1)中,采用Phusion DNA聚合酶(M0530S,NEB)、FastPfu DNA聚合酶(AS221-01,全式金)或Vent DNA聚合酶等高保真DNA聚合酶进行扩增。Further, in step (1), high-fidelity DNA polymerases such as Phusion DNA polymerase (M0530S, NEB), FastPfu DNA polymerase (AS221-01, full gold) or Vent DNA polymerase are used for amplification.
进一步地,在步骤(1)中,采用AxyPrep PCR纯化试剂盒、OMEGA胶回收试剂盒等DNA纯化试剂盒进行纯化。Further, in step (1), DNA purification kits such as AxyPrep PCR purification kit and OMEGA gel recovery kit are used for purification.
进一步地,在步骤(2)中,利用网址http://crispr.mit.edu:8079/gRNA生物信息学网站设计第一gRNA或第二gRNA。Further, in step (2), use the website http://crispr.mit.edu:8079/gRNA bioinformatics website to design the first gRNA or the second gRNA.
进一步地,在步骤(3)中,目的细胞为哺乳动物细胞,如小鼠胚胎干细胞、人胚胎干细胞、Hela、293T等细胞。Further, in step (3), the target cells are mammalian cells, such as mouse embryonic stem cells, human embryonic stem cells, Hela, 293T and other cells.
进一步地,在步骤(3)-(7)中,筛选方法为挑取单个菌落并在96孔板中扩增,然后进行基因型鉴定。Further, in steps (3)-(7), the screening method is to pick a single colony and amplify it in a 96-well plate, and then perform genotype identification.
进一步地,在步骤(1)、(3)、(4)、(5)和步骤(6)中,抗生素为新霉素、潮霉素或嘌呤霉素。步骤(3)中的抗生素与LoxP-抗生素-LoxP序列中的抗生素一致,步骤(5)中的抗生素和FRT-抗生素-FRT-LoxP中的抗生素一致。Further, in step (1), (3), (4), (5) and step (6), the antibiotic is neomycin, hygromycin or puromycin. The antibiotic in step (3) is identical to the antibiotic in the LoxP-antibiotic-LoxP sequence, and the antibiotic in step (5) is identical to the antibiotic in FRT-antibiotic-FRT-LoxP.
进一步地,新霉素(G418)的浓度为200μg/mL-300μg/mL。Further, the concentration of neomycin (G418) is 200 μg/mL-300 μg/mL.
进一步地,潮霉素的浓度为50-1000μg/ml。Further, the concentration of hygromycin is 50-1000 μg/ml.
进一步地,嘌呤霉素的浓度为1-10μg/ml。Further, the concentration of puromycin is 1-10 μg/ml.
进一步地,在步骤(6)中,阿霉素的浓度为0.8μg/mL-1.2μg/mL。Further, in step (6), the concentration of doxorubicin is 0.8 μg/mL-1.2 μg/mL.
进一步地,在步骤(4)和步骤(7)中,4-羟基他莫昔芬的浓度为0.8μM-1.2μM。Further, in step (4) and step (7), the concentration of 4-hydroxytamoxifen is 0.8 μM-1.2 μM.
借由上述方案,本发明至少具有以下优点:By means of the above scheme, the present invention has at least the following advantages:
本发明利用CRISPR/Cas9技术介导同源重组,将重组酶识别位点插入到基因组区域,提高了基因重组效率,节省了时间和人力成本。The invention utilizes CRISPR/Cas9 technology to mediate homologous recombination, inserts the recombinase recognition site into the genome region, improves gene recombination efficiency, and saves time and labor cost.
本发明解决了传统的Cre/loxP系统在基因敲出过程中同源臂长,发生同源重组效率低,基因条件性敲除效率低下的缺点。通过CRISPR/Cas9介导的同源定向修复,只需要设计较短的同源臂便可以在目的基因组区域周围插入loxP位点元件,通过简单的方法便可以产生几个基因的条件性敲除克隆。The invention solves the shortcomings of the traditional Cre/loxP system with long homology arms, low efficiency of homologous recombination and low efficiency of conditional gene knockout in the process of gene knockout. Through CRISPR/Cas9-mediated homology-directed repair, only short homology arms can be designed to insert loxP site elements around the target genomic region, and conditional knockout clones of several genes can be generated by a simple method .
本发明解决了传统条件性敲除在人胚胎干细胞等细胞中重组效率低下的问题。CRISPR/Cas9技术在人胚胎干细胞中能介导较高的同源重组效率,因此极大的提高了LoxP位点靶向导入敲除区域的成功率,为研究在人胚胎干细胞中关键基因的调控功能呢提供了便利条件。The invention solves the problem of low recombination efficiency in cells such as human embryonic stem cells by traditional conditional knockout. CRISPR/Cas9 technology can mediate high homologous recombination efficiency in human embryonic stem cells, thus greatly improving the success rate of targeted introduction of the LoxP site into the knockout region, for the study of the regulation of key genes in human embryonic stem cells. Functions provide convenience.
上述说明仅是本发明技术方案的概述,为了能够更清楚了解本发明的技术手段,并可依照说明书的内容予以实施,以下以本发明的较佳实施例并配合附图详细说明如后。The above description is only an overview of the technical solution of the present invention. In order to understand the technical means of the present invention more clearly, and implement it according to the content of the description, the preferred embodiments of the present invention are described in detail below with the accompanying drawings.
附图说明Description of drawings
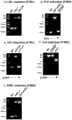
图1是本发明利用CRISPR/Cas9系统构建的基因条件性敲除方法的流程示意图(抗生素使用新霉素);Fig. 1 is the flow chart of the gene conditional knockout method that the present invention utilizes CRISPR/Cas9 system to construct (antibiotic uses neomycin);
图2是本发明实施例1和2所用的打靶载体LNL和FNFL的图谱;Fig. 2 is the map of the used targeting carrier LNL and FNFL of the embodiment of the
图3是本发明实施例1中Eed基因敲除流程图;Fig. 3 is theEed gene knockout flow chart in the embodiment of the
图4是本发明实施例1中Eed敲除克隆的基因型鉴定结果;Fig. 4 is the genotype identification result of Eed knockout clone in the embodiment of the
图5是本发明实施例1中Eed敲除克隆的测序结果;Figure 5 is the sequencing result of the Eed knockout clone in Example 1 of the present invention;
图6是本发明实施例2中SRCAP敲除克隆测序结果。Figure 6 is the sequencing result of the SRCAP knockout clone in Example 2 of the present invention.
具体实施方式Detailed ways
下面结合附图和实施例,对本发明的具体实施方式作进一步详细描述。以下实施例用于说明本发明,但不用来限制本发明的范围。The specific embodiments of the present invention will be described in further detail below with reference to the accompanying drawings and embodiments. The following examples are intended to illustrate the present invention, but not to limit the scope of the present invention.
本发明以下实施例中,LoxP-抗生素-LoxP序列中和FRT-抗生素-FRT-LoxP中的抗生素均选择新霉素,因此分别简称为LNL和FNFL。In the following examples of the present invention, the antibiotics in the LoxP-antibiotic-LoxP sequence and FRT-antibiotic-FRT-LoxP are both selected as neomycin, so they are abbreviated as LNL and FNFL respectively.
实施例1Example 1
本实施例提供了一种选择性敲除小鼠胚胎干细胞(目的细胞)中Eed基因(目的基因)的方法,具体包括以下步骤:The present embodiment provides a method for selectively knocking out the Eed gene (target gene) in mouse embryonic stem cells (target cells), which specifically includes the following steps:
(1)构建Eed基因左侧打靶质粒:在Eed基因2号外显子上游适当位置选取LNL序列待插入位点,该位点上下游各选取500bp至1kb范围的左右同源臂,使用高保真Phusion DNA聚合酶(M0530S,NEB)扩增LNL序列待插入位点左右同源臂,然后用AxyPrep PCR纯化试剂盒纯化,左臂利用KpnⅠ和EcoRⅠ限制性内切酶消化PCR产物,右臂用BamHⅠ和SacⅡ消化,插入到含有LNL序列的质粒中,构建目的基因左侧打靶质粒。(1) Construction of a targeting plasmid on the left side of the Eed gene: Select the LNL sequence to be inserted at the appropriate position upstream of
(2)利用网址http://crispr.mit.edu:8079/gRNA生物信息学网站设计目的基因左侧打靶质粒相对应的gRNA,其核苷酸序列如SEQ ID No.1-SEQ ID No.2所示。Eed-LNLtargeting gRNA-1:GGCTTCACTCTAACGACTGT(SEQ ID No.1),Eed-LNL targeting gRNA-2:AGTCGTTAGAGTGAAGCCCA(SEQ ID No.2)。具体构建方法如下:首先按照保护碱基、识别位点、酶切位点、sgRNA和U6启动子引物的顺序合成新的引物,接着以PGL3载体为模板,扩增出含有U6启动子和sgRNA的目的基因。最后将PCR产物纯化酶切后与同样酶切过的PGL3-U6-2sgRNA-puro载体连接即可获得gRNA表达载体。(2) Using the website http://crispr.mit.edu:8079/gRNA bioinformatics website to design the gRNA corresponding to the targeting plasmid on the left side of the target gene, the nucleotide sequence of which is as shown in SEQ ID No.1-SEQ ID No. 2 shown. Eed-LNL targeting gRNA-1: GGCTTCACTCTAACGACTGT (SEQ ID No. 1), Eed-LNL targeting gRNA-2: AGTCGTTAGAGTGAAGCCCA (SEQ ID No. 2). The specific construction method is as follows: First, synthesize new primers in the order of protected bases, recognition sites, restriction sites, sgRNA and U6 promoter primers, and then use the PGL3 vector as a template to amplify the primers containing U6 promoter and sgRNA. target gene. Finally, the PCR product was purified and digested and then connected with the same digested PGL3-U6-2sgRNA-puro vector to obtain the gRNA expression vector.
(3)Eed左侧打靶质粒整合基因组中:将Cas9质粒,步骤(2)构建的gRNA的表达质粒和目的基因左侧打靶质粒同时转入到目的细胞中,转染后48小时用浓度为250μg/mL G418进行细胞选择。在G418选择6天后,挑取单个菌落并在96孔板中扩增用于随后的基因分型,选择成功插入了目的基因左侧打靶质粒的细胞。胚胎干细胞培养条件为:37℃,5%二氧化碳的培养箱中培养(3) The Eed left-side targeting plasmid is integrated into the genome: the Cas9 plasmid, the gRNA expression plasmid constructed in step (2) and the left-side targeting plasmid of the target gene are simultaneously transferred into the target cells, at a concentration of 250 μg 48 hours after transfection /mL G418 for cell selection. After 6 days of G418 selection, single colonies were picked and expanded in 96-well plates for subsequent genotyping, selecting cells that successfully inserted the targeting plasmid to the left of the gene of interest. Embryonic stem cell culture conditions are: 37 ° C, 5% carbon dioxide incubator
(4)Cre酶介导的细胞同源重组:用浓度为1μM的4-OHT(H7904,Sigma)处理成功插入了目的基因左侧打靶质粒的细胞5天,诱导Cre-LoxP介导的同源重组,挑取单独的菌落进行PCR基因分型,确认目的基因左侧成功插入loxp位点。(4) Cell homologous recombination mediated by Cre enzyme: The cells successfully inserted with the targeting plasmid on the left side of the target gene were treated with 4-OHT (H7904, Sigma) at a concentration of 1 μM for 5 days to induce Cre-LoxP-mediated homologous recombination. After recombination, individual colonies were picked for PCR genotyping to confirm the successful insertion of the loxp site on the left side of the target gene.
(5)构建Eed右侧打靶质粒:在目的基因右侧适当位置选取FNFL序列待插入位点,该位点上下游各选取500bp至1kb范围的左右同源臂,使用高保真Phusion DNA聚合酶(M0530S,NEB)扩增FNFL序列待插入位点左右同源臂,然后用AxyPrep PCR纯化试剂盒纯化,左臂利用KpnⅠ和EcoRⅠ限制性内切酶消化PCR产物,右臂用BamHⅠ和SacⅡ消化,插入到含有FNFL序列的质粒中,构建Eed右侧打靶质粒。(5) construct the Eed right side targeting plasmid: select the FNFL sequence to be inserted at the appropriate position on the right side of the target gene, select the left and right homology arms in the range of 500bp to 1kb at the upstream and downstream of the site, and use high-fidelity Phusion DNA polymerase ( M0530S, NEB) to amplify the left and right homology arms of the FNFL sequence to be inserted at the site to be inserted, and then purified with AxyPrep PCR purification kit. The left arm was digested with KpnI and EcoRI restriction endonucleases, and the right arm was digested with BamHI and SacII. Into the plasmid containing the FNFL sequence, construct the Eed right-side targeting plasmid.
(6)利用网址http://crispr.mit.edu:8079/gRNA生物信息学网站设计目的基因右侧打靶质粒相对应的gRNA,其核苷酸序列如SEQ ID No.3-SEQ ID No.4所示。为Eed-FNFLtargeting gRNA-1:TGTGTAAAGGCCGGCTTACC(SEQ ID No.3);Eed-FNFL targeting gRNA-2:AGGCCGGCTTACCAGGCACG(SEQ ID No.4)。(6) Use the website http://crispr.mit.edu:8079/gRNA bioinformatics website to design the gRNA corresponding to the targeting plasmid on the right side of the target gene, the nucleotide sequence of which is as shown in SEQ ID No.3-SEQ ID No. 4 shown. Eed-FNFL targeting gRNA-1: TGTGTAAAGGCCGGCTTACC (SEQ ID No. 3); Eed-FNFL targeting gRNA-2: AGGCCGGCTTACCAGGCACG (SEQ ID No. 4).
(7)Eed右侧打靶质粒整合基因组中:将Cas9质粒,步骤(6)构建的gRNA的表达质粒和Eed右侧打靶质粒同时转入到目的基因左侧成功插入loxp位点的目的细胞中,转染后48小时用浓度为250μg/mL的G418进行细胞选择。在G418选择6天后,挑取单个菌落并在96孔板中扩增用于随后的基因分型,选择成功插入了目的基因左侧和右侧打靶质粒的细胞。(7) The targeting plasmid on the right side of Eed is integrated into the genome: the Cas9 plasmid, the gRNA expression plasmid constructed in step (6), and the targeting plasmid on the right side of Eed are simultaneously transferred into the target cell where the loxp site is successfully inserted on the left side of the target gene, Cell selection was performed 48 hours after transfection with G418 at a concentration of 250 μg/mL. After 6 days of G418 selection, single colonies were picked and expanded in 96-well plates for subsequent genotyping, selecting cells with successfully inserted targeting plasmids to the left and right of the gene of interest.
(8)FLIP酶介导的细胞同源重组:用强力霉素(doxycycline)处理步骤(7)中成功插入了目的基因左侧和右侧打靶质粒的细胞5天,诱导FLP-FRT介导的同源重组,挑取单独的菌落进行PCR基因型鉴定,确认Eed基因左侧和右侧均成功插入loxp位点。(8) Homologous recombination mediated by FLIP enzyme: cells into which targeting plasmids on the left and right sides of the target gene were successfully inserted in step (7) were treated with doxycycline for 5 days to induce FLP-FRT-mediated recombination. Homologous recombination, picking individual colonies for PCR genotype identification, confirming that both the left and right sides of the Eed gene were successfully inserted into the loxp site.
(9)Cre酶介导的细胞同源重组:用浓度为1μM的4-OHT处理目的基因左右侧成功插入loxP位点的细胞5天,诱导Cre-LoxP介导的同源重组,挑取单独的菌落进行PCR基因分型,成功敲除了小鼠胚胎干细胞中的Eed基因。(9) Homologous recombination mediated by Cre enzyme: Treat the cells successfully inserted into the loxP site on the left and right sides of the target gene with 4-OHT at a concentration of 1 μM for 5 days to induce Cre-LoxP-mediated homologous recombination. PCR genotyping of the colonies successfully knocked out the Eed gene in mouse embryonic stem cells.
实施例2Example 2
本实施例提供了一种条件性敲除小鼠胚胎干细胞(目的细胞)中SRCAP基因(目的基因)的方法,具体包括以下步骤:The present embodiment provides a method for conditionally knocking out the SRCAP gene (target gene) in mouse embryonic stem cells (target cells), which specifically includes the following steps:
(1)构建SRCAP基因敲除区域左侧LNL打靶质粒:选取SRCAP基因的5号-8号外显子作为目的敲除区域,以5号外显子上游某位置作为LNL插入位点。首先以小鼠基因组为模板,利用Pusion高保真DNA聚合酶在插入位点的上下游扩增出500bp至1kb大小的左右同源臂,而后用AxyPrep PCR纯化试剂盒纯化;接着用KpnⅠ和EcoRⅠ限制性内切酶消化左臂PCR纯化产物,右臂则用BamHⅠ和SacⅡ消化,之后将两个同源臂依次连接到LNL载体上得到靶向目的敲除区域左侧的LNL质粒。(1) Construction of the LNL targeting plasmid on the left side of the SRCAP gene knockout region: Select exons 5-8 of the SRCAP gene as the target knockout region, and a position upstream of
(2)利用网址http://crispr.mit.edu:8079/gRNA生物信息学网站设计SRCAP基因目的敲除区域左侧打靶质粒相对应的gRNA-1:AGGTAGTATCCCTAAGTAGT(SEQ ID No.5),sgRNA-2:ATCTCTTAGCCAACTACTTA(SEQ ID No.6)。(2) Using the website http://crispr.mit.edu:8079/gRNA bioinformatics to design gRNA-1 corresponding to the targeting plasmid on the left side of the target knockout region of the SRCAP gene: AGGTAGTATCCCTAAGTAGT (SEQ ID No. 5), sgRNA -2: ATCTCTTAGCCAACTACTTA (SEQ ID No. 6).
(3)目的敲除区域左侧打靶质粒LNL盒整合至基因组:将Cas9质粒,步骤(2)构建的gRNA的表达质粒和LNL靶向质粒同时电转到目的细胞中,转染48小时后用浓度为250μg/mL的G418进行筛选,将胚胎干细胞于37℃且含有5%二氧化碳的培养箱中培养,并保持一定湿度。G418选择6天后,挑取单个菌落并在96孔板中扩增用于随后的基因型鉴定,并获得基因组中双等位基因靶向导入LNL序列的细胞。(3) Integration of the targeting plasmid LNL cassette on the left side of the target knockout region into the genome: The Cas9 plasmid, the gRNA expression plasmid constructed in step (2) and the LNL targeting plasmid were electroporated into the target cells at the same time, and 48 hours after transfection, the concentration of For screening at 250 μg/mL of G418, embryonic stem cells were cultured at 37°C in an incubator containing 5% carbon dioxide and maintained at a certain humidity. Six days after G418 selection, single colonies were picked and expanded in 96-well plates for subsequent genotyping, and cells with biallelic targeted introduction of LNL sequences in the genome were obtained.
(4)Cre酶介导的细胞同源重组:用浓度为1μM的4-OHT(Sigma,H7904)处理步骤(3)中筛选获得的细胞5天,让其于37℃且含有5%二氧化碳的培养箱中培养,并保持一定湿度,以诱导Cre-LoxP介导的同源重组,挑取单菌落进行PCR基因型鉴定,确认细胞基因组上LoxP双等位基因插入。(4) Cre-mediated homologous recombination of cells: The cells obtained in step (3) were treated with 4-OHT (Sigma, H7904) at a concentration of 1 μM for 5 days, and allowed to cool at 37° C. in a 5% carbon dioxide-containing environment. Culture in an incubator and maintain a certain humidity to induce Cre-LoxP-mediated homologous recombination, pick a single colony for PCR genotype identification, and confirm the LoxP biallelic insertion on the cell genome.
(5)抗neomycin捕获盒的药耐性缺失:将步骤(4)获得的LNL捕获盒缺失的细胞扩增两份,一份加入浓度为250μg/mL的G418处理3天,另一份不作加药处理。观察到加药处理的细胞死亡,确认细胞中的LNL捕获盒缺失。不加药的细胞用作后续实验。(5) Lack of drug resistance of the anti-neomycin capture cassette: the cells with the missing LNL capture cassette obtained in step (4) were expanded in two copies, one was treated with G418 at a concentration of 250 μg/mL for 3 days, and the other was not added deal with. Drug-treated cell death was observed, confirming the absence of the LNL capture cassette in the cells. Unmedicated cells were used for subsequent experiments.
(6)构建目的敲除区域右侧FNFL打靶质粒:在目的基因SRCAP的8号外显子下游位置选取FNFL序列待插入位点。首先以小鼠基因组为模板,利用Pusion高保真DNA聚合酶(M0530S,NEB)在插入位点的上下游扩增出500bp至1kb大小的左右同源臂,而后用AxyPrepPCR纯化试剂盒纯化;接着利用KpnⅠ和EcoRⅠ限制性内切酶消化左臂PCR纯化产物,右臂则用BamHⅠ和SacⅡ消化,之后将两个同源臂依次连接到FNFL载体上得到靶向SRCAP目的敲除区域右侧的FNFL质粒。(6) Constructing the FNFL targeting plasmid on the right side of the target knockout region: select the FNFL sequence to be inserted at the downstream position of exon 8 of the target gene SRCAP. First, using the mouse genome as a template, using Pusion high-fidelity DNA polymerase (M0530S, NEB) to amplify the left and right homology arms of 500bp to 1kb in size at the upstream and downstream of the insertion site, and then use the AxyPrepPCR purification kit to purify; The PCR-purified product of the left arm was digested with KpnI and EcoRI restriction enzymes, and the right arm was digested with BamHI and SacII. After that, the two homology arms were sequentially ligated to the FNFL vector to obtain the FNFL plasmid targeting the right side of the SRCAP target knockout region .
(7)利用网址http://crispr.mit.edu:8079/gRNA生物信息学网站设计SRCAP基因目的敲除区域右侧打靶质粒相对应的gRNA-1:GAGCTCAAGCCGCAGTTCGC(SEQ ID No.7),gRNA-2:AGTGGGGATCCTGCGAACTG(SEQ ID No.8),从而构建sgRNA表达质粒。(7) Using the website http://crispr.mit.edu:8079/gRNA bioinformatics to design the gRNA-1 corresponding to the targeting plasmid on the right side of the target knockout region of the SRCAP gene: GAGCTCAAGCCGCAGTTCGC (SEQ ID No. 7), gRNA -2: AGTGGGGATCCTGCGAACTG (SEQ ID No. 8), thereby constructing an sgRNA expression plasmid.
(8)目的敲除区域右侧打靶质粒FNFL整合至基因组:将Cas9质粒,步骤(7)构建的gRNA表达质粒和步骤(6)构建的FNFL打靶质粒同时转入到待敲除区域左侧成功插入LoxP位点的目的细胞中,转染后48小时后用浓度为250μg/mL的G418进行细胞筛选。在G418选择6天后,挑取单个菌落并在96孔板中扩增用于随后的基因型鉴定,选择基因组上成功插入了右侧打靶质粒FNFL序列的细胞。(8) Integration of the targeting plasmid FNFL on the right side of the target knockout region into the genome: The Cas9 plasmid, the gRNA expression plasmid constructed in step (7) and the FNFL targeting plasmid constructed in step (6) were simultaneously transferred to the left side of the region to be knocked out. The cells of interest inserted into the LoxP site were screened with G418 at a concentration of 250 μg/
(9)FLIP酶介导的细胞同源重组:用强力霉素(doxycycline)处理步骤(8)中目的敲除区域右侧成功插入FNFL序列的细胞5天,诱导FLP-FRT介导的同源重组,挑取单独的菌落进行PCR基因型鉴定,确认FNFL捕获盒缺失剩一个FRT位点和一个LoxP位点。至此,目的敲除区域(SRCAP基因5号外显子)基因组上下游分别含有一个LoxP位点。(9) Homologous recombination mediated by FLIP enzyme: The cells in which the FNFL sequence was successfully inserted to the right of the target knockout region in step (8) were treated with doxycycline for 5 days to induce FLP-FRT-mediated homologous recombination. After recombination, individual colonies were picked for PCR genotype identification, and it was confirmed that the FNFL capture cassette was missing one FRT site and one LoxP site. So far, the target knockout region (SRCAP gene exon 5) contains a LoxP site in the upstream and downstream of the genome.
(10)Cre酶介导的细胞同源重组:用浓度为1μM的4-OHT处理步骤(9)中获得的目的敲除区域左右侧各有一个LoxP位点的细胞5天,诱导Cre-LoxP介导的同源重组,挑取单独的菌落进行PCR基因型鉴定,确认成功敲除了小鼠胚胎干细胞中SRCAP基因的5号-8号外显子。(10) Cre-mediated homologous recombination: cells with a LoxP site on the left and right sides of the target knockout region obtained in step (9) were treated with 4-OHT at a concentration of 1 μM for 5 days to induce Cre-LoxP Mediated homologous recombination, picking individual colonies for PCR genotype identification, confirming the successful knockout of exons 5-8 of the SRCAP gene in mouse embryonic stem cells.
以上所述仅是本发明的优选实施方式,并不用于限制本发明,应当指出,对于本技术领域的普通技术人员来说,在不脱离本发明技术原理的前提下,还可以做出若干改进和变型,这些改进和变型也应视为本发明的保护范围。The above are only the preferred embodiments of the present invention and are not intended to limit the present invention. It should be pointed out that for those skilled in the art, some improvements can be made without departing from the technical principles of the present invention. These improvements and modifications should also be regarded as the protection scope of the present invention.



序列表sequence listing
<110> 苏州大学<110> Soochow University
<120> 利用CRISPR/Cas9系统构建的基因条件性敲除方法<120> Conditional gene knockout method constructed by CRISPR/Cas9 system
<160> 8<160> 8
<170> SIPOSequenceListing 1.0<170> SIPOSequenceListing 1.0
<210> 1<210> 1
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 1<400> 1
ggcttcactc taacgactgt 20ggcttcactc taacgactgt 20
<210> 2<210> 2
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 2<400> 2
agtcgttaga gtgaagccca 20
<210> 3<210> 3
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 3<400> 3
tgtgtaaagg ccggcttacc 20
<210> 4<210> 4
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 4<400> 4
aggccggctt accaggcacg 20
<210> 5<210> 5
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 5<400> 5
aggtagtatc cctaagtagt 20aggtagtatc cctaagtagt 20
<210> 6<210> 6
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 6<400> 6
atctcttagc caactactta 20
<210> 7<210> 7
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 7<400> 7
gagctcaagc cgcagttcgc 20
<210> 8<210> 8
<211> 20<211> 20
<212> DNA<212> DNA
<213> (人工序列)<213> (artificial sequence)
<400> 8<400> 8
agtggggatc ctgcgaactg 20agtggggatc ctgcgaactg 20
Claims (9)
Priority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201810300747.7ACN108441520B (en) | 2018-04-04 | 2018-04-04 | Conditional gene knockout method constructed by CRISPR/Cas9 system |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201810300747.7ACN108441520B (en) | 2018-04-04 | 2018-04-04 | Conditional gene knockout method constructed by CRISPR/Cas9 system |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN108441520A CN108441520A (en) | 2018-08-24 |
| CN108441520Btrue CN108441520B (en) | 2020-07-31 |
Family
ID=63198326
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201810300747.7AActiveCN108441520B (en) | 2018-04-04 | 2018-04-04 | Conditional gene knockout method constructed by CRISPR/Cas9 system |
Country Status (1)
| Country | Link |
|---|---|
| CN (1) | CN108441520B (en) |
Families Citing this family (33)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| EP3613852A3 (en) | 2011-07-22 | 2020-04-22 | President and Fellows of Harvard College | Evaluation and improvement of nuclease cleavage specificity |
| US20150044192A1 (en) | 2013-08-09 | 2015-02-12 | President And Fellows Of Harvard College | Methods for identifying a target site of a cas9 nuclease |
| US9359599B2 (en) | 2013-08-22 | 2016-06-07 | President And Fellows Of Harvard College | Engineered transcription activator-like effector (TALE) domains and uses thereof |
| US9228207B2 (en) | 2013-09-06 | 2016-01-05 | President And Fellows Of Harvard College | Switchable gRNAs comprising aptamers |
| US9526784B2 (en) | 2013-09-06 | 2016-12-27 | President And Fellows Of Harvard College | Delivery system for functional nucleases |
| US9322037B2 (en) | 2013-09-06 | 2016-04-26 | President And Fellows Of Harvard College | Cas9-FokI fusion proteins and uses thereof |
| US11053481B2 (en) | 2013-12-12 | 2021-07-06 | President And Fellows Of Harvard College | Fusions of Cas9 domains and nucleic acid-editing domains |
| EP3177718B1 (en) | 2014-07-30 | 2022-03-16 | President and Fellows of Harvard College | Cas9 proteins including ligand-dependent inteins |
| SG10202104041PA (en) | 2015-10-23 | 2021-06-29 | Harvard College | Nucleobase editors and uses thereof |
| WO2018027078A1 (en) | 2016-08-03 | 2018-02-08 | President And Fellows Of Harard College | Adenosine nucleobase editors and uses thereof |
| WO2018031683A1 (en) | 2016-08-09 | 2018-02-15 | President And Fellows Of Harvard College | Programmable cas9-recombinase fusion proteins and uses thereof |
| WO2018039438A1 (en) | 2016-08-24 | 2018-03-01 | President And Fellows Of Harvard College | Incorporation of unnatural amino acids into proteins using base editing |
| EP3526320A1 (en) | 2016-10-14 | 2019-08-21 | President and Fellows of Harvard College | Aav delivery of nucleobase editors |
| US10745677B2 (en) | 2016-12-23 | 2020-08-18 | President And Fellows Of Harvard College | Editing of CCR5 receptor gene to protect against HIV infection |
| EP3592853A1 (en) | 2017-03-09 | 2020-01-15 | President and Fellows of Harvard College | Suppression of pain by gene editing |
| EP3592381A1 (en) | 2017-03-09 | 2020-01-15 | President and Fellows of Harvard College | Cancer vaccine |
| JP2020510439A (en) | 2017-03-10 | 2020-04-09 | プレジデント アンド フェローズ オブ ハーバード カレッジ | Base-editing factor from cytosine to guanine |
| WO2018176009A1 (en) | 2017-03-23 | 2018-09-27 | President And Fellows Of Harvard College | Nucleobase editors comprising nucleic acid programmable dna binding proteins |
| WO2018209320A1 (en) | 2017-05-12 | 2018-11-15 | President And Fellows Of Harvard College | Aptazyme-embedded guide rnas for use with crispr-cas9 in genome editing and transcriptional activation |
| CN111801345A (en) | 2017-07-28 | 2020-10-20 | 哈佛大学的校长及成员们 | Methods and compositions using an evolved base editor for Phage Assisted Continuous Evolution (PACE) |
| WO2019139645A2 (en) | 2017-08-30 | 2019-07-18 | President And Fellows Of Harvard College | High efficiency base editors comprising gam |
| CA3082251A1 (en) | 2017-10-16 | 2019-04-25 | The Broad Institute, Inc. | Uses of adenosine base editors |
| EP3724214A4 (en) | 2017-12-15 | 2021-09-01 | The Broad Institute Inc. | SYSTEMS AND PROCEDURES FOR PREDICTING REPAIR RESULTS IN GENE ENGINEERING |
| US12157760B2 (en) | 2018-05-23 | 2024-12-03 | The Broad Institute, Inc. | Base editors and uses thereof |
| WO2020072480A1 (en)* | 2018-10-01 | 2020-04-09 | Lonza Ltd | Ssi cells with predictable and stable transgene expression and methods of formation |
| US12281338B2 (en) | 2018-10-29 | 2025-04-22 | The Broad Institute, Inc. | Nucleobase editors comprising GeoCas9 and uses thereof |
| US12351837B2 (en) | 2019-01-23 | 2025-07-08 | The Broad Institute, Inc. | Supernegatively charged proteins and uses thereof |
| WO2020191246A1 (en) | 2019-03-19 | 2020-09-24 | The Broad Institute, Inc. | Methods and compositions for editing nucleotide sequences |
| WO2021072328A1 (en) | 2019-10-10 | 2021-04-15 | The Broad Institute, Inc. | Methods and compositions for prime editing rna |
| AU2021267940A1 (en) | 2020-05-08 | 2022-12-08 | President And Fellows Of Harvard College | Methods and compositions for simultaneous editing of both strands of a target double-stranded nucleotide sequence |
| CN113151354B (en)* | 2021-03-22 | 2022-03-08 | 中国农业科学院兰州兽医研究所 | Vector for conditional knockout of target gene and method for conditional knockout of target gene |
| CN116987733A (en)* | 2023-04-24 | 2023-11-03 | 广西医科大学 | Vector for targeted knockout of VASN gene and application thereof |
| CN117070434A (en)* | 2023-08-31 | 2023-11-17 | 西南医科大学 | A live attenuated vaccine strain with slyA gene deletion of Klebsiella pneumoniae and its preparation method and application |
Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN104726494A (en)* | 2015-02-12 | 2015-06-24 | 中国人民解放军第二军医大学 | Method for constructing chromosome translocation stem cell and animal model by CRISPR-Cas9 technology |
| CA2963080A1 (en)* | 2014-10-01 | 2016-04-07 | The General Hospital Corporation | Methods for increasing efficiency of nuclease-induced homology-directed repair |
| CN105637087A (en)* | 2013-09-18 | 2016-06-01 | 科马布有限公司 | Methods, cells and organisms |
| CN105985985A (en)* | 2016-05-06 | 2016-10-05 | 苏州大学 | Preparation method of allogeneic mesenchymal stem cells edited by CRISPR technology and optimized by IGF (insulin-like growth factor) and application of allogeneic mesenchymal stem cells in treatment of myocardial infarction |
- 2018
- 2018-04-04CNCN201810300747.7Apatent/CN108441520B/enactiveActive
Patent Citations (4)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN105637087A (en)* | 2013-09-18 | 2016-06-01 | 科马布有限公司 | Methods, cells and organisms |
| CA2963080A1 (en)* | 2014-10-01 | 2016-04-07 | The General Hospital Corporation | Methods for increasing efficiency of nuclease-induced homology-directed repair |
| CN104726494A (en)* | 2015-02-12 | 2015-06-24 | 中国人民解放军第二军医大学 | Method for constructing chromosome translocation stem cell and animal model by CRISPR-Cas9 technology |
| CN105985985A (en)* | 2016-05-06 | 2016-10-05 | 苏州大学 | Preparation method of allogeneic mesenchymal stem cells edited by CRISPR technology and optimized by IGF (insulin-like growth factor) and application of allogeneic mesenchymal stem cells in treatment of myocardial infarction |
Non-Patent Citations (1)
| Title |
|---|
| One-step generation of conditional and reversible gene knockouts;Amanda Andersson-Rolf等;《Nat Methods》;20170130;第14卷(第3期);摘要,简介,方法部分* |
Also Published As
| Publication number | Publication date |
|---|---|
| CN108441520A (en) | 2018-08-24 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN108441520B (en) | Conditional gene knockout method constructed by CRISPR/Cas9 system | |
| US20180355382A1 (en) | Large genomic dna knock-in and uses thereof | |
| CN103388006B (en) | A kind of construction process of site-directed point mutation | |
| Boroviak et al. | Chromosome engineering in zygotes with CRISPR/C as9 | |
| CN106047930B (en) | Preparation method of Flox rat with conditional knockout of PS1 gene | |
| Shao et al. | CRISPR/Cas-mediated genome editing in the rat via direct injection of one-cell embryos | |
| Wefers et al. | Gene editing in mouse zygotes using the CRISPR/Cas9 system | |
| EP3546575B1 (en) | Genome editing method | |
| Park et al. | Targeted gene knock-in by CRISPR/Cas ribonucleoproteins in porcine zygotes | |
| CN110551759B (en) | Composition and method for improving recombination efficiency of transgenic cells | |
| CN106467909A (en) | A kind of method that acquisition glyphosate-resistant rice is replaced by nucleotide fixed point | |
| TR201816074T4 (en) | Methods and compositions for targeted genetic modifications and methods of use. | |
| EP2858486A2 (en) | Methods and compositions for generating conditional knock-out alleles | |
| US20190169653A1 (en) | Method for preparing gene knock-in cells | |
| CN110643636B (en) | A kind of method and application of knocking out MSTNa&b gene of bream | |
| Li-Fang et al. | Generation of genetically modified mice using CRISPR/Cas9 and haploid embryonic stem cell systems | |
| Ge et al. | Efficient homologous recombination in mice using long single stranded DNA and CRISPR Cas9 nickase | |
| CN106148406B (en) | Porcine ApoE gene knockout vector and its construction method and application | |
| CN105315352B (en) | Insect site-directed gene knock-in composition and application method and application thereof | |
| CN114934073A (en) | Construction methods and applications of hoxa1a knockout zebrafish mutants | |
| CN111549070B (en) | Methods of editing multiple copies of X chromosome genes to achieve animal sex control | |
| CN108866102A (en) | A kind of construction method of Adgrv1 gene Y6236fsX1 mutant animals model | |
| CN109628447B (en) | sgRNA specifically targeting sheep-friendly site H11 and its coding DNA and application | |
| CN111793654B (en) | A method for improving the efficiency of CRISPR/Cas9-mediated biallelic mutation and its application | |
| Yu et al. | Application of genome editing technology to microrna research in mammalians |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| GR01 | Patent grant | ||
| GR01 | Patent grant |