CN106831967B - Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus - Google Patents
Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virusDownload PDFInfo
- Publication number
- CN106831967B CN106831967BCN201510882608.6ACN201510882608ACN106831967BCN 106831967 BCN106831967 BCN 106831967BCN 201510882608 ACN201510882608 ACN 201510882608ACN 106831967 BCN106831967 BCN 106831967B
- Authority
- CN
- China
- Prior art keywords
- iaa10
- protein
- rice
- rdv
- sequence
- Prior art date
- Legal status (The legal status is an assumption and is not a legal conclusion. Google has not performed a legal analysis and makes no representation as to the accuracy of the status listed.)
- Active
Links
- 241000702632Rice dwarf virusSpecies0.000titleclaimsabstractdescription76
- 241000196324EmbryophytaSpecies0.000titleclaimsabstractdescription46
- 101150081360IAA10 geneProteins0.000titleclaimsabstractdescription12
- 230000014509gene expressionEffects0.000titledescription13
- 108090000623proteins and genesProteins0.000claimsabstractdescription142
- 235000007164Oryza sativaNutrition0.000claimsabstractdescription75
- 235000009566riceNutrition0.000claimsabstractdescription74
- 239000013598vectorSubstances0.000claimsabstractdescription53
- 125000003275alpha amino acid groupChemical group0.000claimsabstractdescription13
- 241000209094OryzaSpecies0.000claimsabstract4
- 102000004169proteins and genesHuman genes0.000claimsdescription103
- 230000009368gene silencing by RNAEffects0.000claimsdescription18
- 108091030071RNAIProteins0.000claimsdescription16
- 108020004414DNAProteins0.000claimsdescription11
- 102000053602DNAHuman genes0.000claimsdescription8
- 230000002441reversible effectEffects0.000claimsdescription5
- FWMNVWWHGCHHJJ-SKKKGAJSSA-N4-amino-1-[(2r)-6-amino-2-[[(2r)-2-[[(2r)-2-[[(2r)-2-amino-3-phenylpropanoyl]amino]-3-phenylpropanoyl]amino]-4-methylpentanoyl]amino]hexanoyl]piperidine-4-carboxylic acidChemical compoundC([C@H](C(=O)N[C@H](CC(C)C)C(=O)N[C@H](CCCCN)C(=O)N1CCC(N)(CC1)C(O)=O)NC(=O)[C@H](N)CC=1C=CC=CC=1)C1=CC=CC=C1FWMNVWWHGCHHJJ-SKKKGAJSSA-N0.000claimsdescription4
- 230000000295complement effectEffects0.000claimsdescription3
- 230000009261transgenic effectEffects0.000abstractdescription27
- 201000010099diseaseDiseases0.000abstractdescription15
- 208000037265diseases, disorders, signs and symptomsDiseases0.000abstractdescription15
- 238000002474experimental methodMethods0.000abstractdescription15
- 230000001105regulatory effectEffects0.000abstractdescription7
- 101710132601Capsid proteinProteins0.000abstractdescription3
- 101710094648Coat proteinProteins0.000abstractdescription3
- 102100021181Golgi phosphoprotein 3Human genes0.000abstractdescription3
- 101710125418Major capsid proteinProteins0.000abstractdescription3
- 101710141454NucleoproteinProteins0.000abstractdescription3
- 101710083689Probable capsid proteinProteins0.000abstractdescription3
- 238000009825accumulationMethods0.000abstractdescription2
- 235000018102proteinsNutrition0.000description100
- 240000007594Oryza sativaSpecies0.000description76
- SEOVTRFCIGRIMH-UHFFFAOYSA-Nindole-3-acetic acidChemical compoundC1=CC=C2C(CC(=O)O)=CNC2=C1SEOVTRFCIGRIMH-UHFFFAOYSA-N0.000description40
- 239000000047productSubstances0.000description34
- 238000000034methodMethods0.000description27
- 239000013612plasmidSubstances0.000description25
- 238000003752polymerase chain reactionMethods0.000description23
- 230000015556catabolic processEffects0.000description19
- 238000006731degradation reactionMethods0.000description19
- 239000000284extractSubstances0.000description16
- 241000208125NicotianaSpecies0.000description15
- 235000002637Nicotiana tabacumNutrition0.000description15
- 229930192334AuxinNatural products0.000description14
- 206010020649HyperkeratosisDiseases0.000description14
- 239000002363auxinSubstances0.000description14
- 108091032973(ribonucleotides)n+mProteins0.000description12
- 239000002609mediumSubstances0.000description12
- 208000024891symptomDiseases0.000description12
- 241000589158AgrobacteriumSpecies0.000description10
- 229930191978GibberellinNatural products0.000description9
- 230000015572biosynthetic processEffects0.000description9
- 238000010276constructionMethods0.000description9
- 239000003448gibberellinSubstances0.000description9
- 239000007788liquidSubstances0.000description9
- IJGRMHOSHXDMSA-UHFFFAOYSA-NAtomic nitrogenChemical compoundN#NIJGRMHOSHXDMSA-UHFFFAOYSA-N0.000description8
- 108010067758ent-kaurene oxidaseProteins0.000description8
- IXORZMNAPKEEDV-OBDJNFEBSA-Ngibberellin A3Chemical classC([C@@]1(O)C(=C)C[C@@]2(C1)[C@H]1C(O)=O)C[C@H]2[C@]2(C=C[C@@H]3O)[C@H]1[C@]3(C)C(=O)O2IXORZMNAPKEEDV-OBDJNFEBSA-N0.000description8
- 230000037039plant physiologyEffects0.000description8
- 230000002829reductive effectEffects0.000description8
- 238000012216screeningMethods0.000description8
- 239000006228supernatantSubstances0.000description8
- 102000004190EnzymesHuman genes0.000description7
- 108090000790EnzymesProteins0.000description7
- 101710088566Flagellar hook-associated protein 2Proteins0.000description7
- 102100034408Nuclear transcription factor Y subunit alphaHuman genes0.000description7
- 239000000872bufferSubstances0.000description7
- 210000004027cellAnatomy0.000description7
- 239000013604expression vectorSubstances0.000description7
- 230000003993interactionEffects0.000description7
- 230000002018overexpressionEffects0.000description7
- 239000000843powderSubstances0.000description7
- 238000012163sequencing techniqueMethods0.000description7
- 238000012546transferMethods0.000description7
- 241001414720CicadellidaeSpecies0.000description6
- PEDCQBHIVMGVHV-UHFFFAOYSA-NGlycerineChemical compoundOCC(O)COPEDCQBHIVMGVHV-UHFFFAOYSA-N0.000description6
- FAPWRFPIFSIZLT-UHFFFAOYSA-MSodium chlorideChemical compound[Na+].[Cl-]FAPWRFPIFSIZLT-UHFFFAOYSA-M0.000description6
- 101150044379TIR1 geneProteins0.000description6
- 230000001580bacterial effectEffects0.000description6
- 238000001727in vivoMethods0.000description6
- 208000015181infectious diseaseDiseases0.000description6
- YBYRMVIVWMBXKQ-UHFFFAOYSA-Nphenylmethanesulfonyl fluorideChemical compoundFS(=O)(=O)CC1=CC=CC=C1YBYRMVIVWMBXKQ-UHFFFAOYSA-N0.000description6
- 238000002415sodium dodecyl sulfate polyacrylamide gel electrophoresisMethods0.000description6
- 238000001262western blotMethods0.000description6
- 240000004808Saccharomyces cerevisiaeSpecies0.000description5
- 239000002299complementary DNASubstances0.000description5
- 238000000338in vitroMethods0.000description5
- 108091008146restriction endonucleasesProteins0.000description5
- 238000010839reverse transcriptionMethods0.000description5
- XLYOFNOQVPJJNP-UHFFFAOYSA-NwaterChemical compoundOXLYOFNOQVPJJNP-UHFFFAOYSA-N0.000description5
- 102100034343IntegraseHuman genes0.000description4
- TWRXJAOTZQYOKJ-UHFFFAOYSA-LMagnesium chlorideChemical compound[Mg+2].[Cl-].[Cl-]TWRXJAOTZQYOKJ-UHFFFAOYSA-L0.000description4
- 108010092799RNA-directed DNA polymeraseProteins0.000description4
- 238000006243chemical reactionMethods0.000description4
- 238000001976enzyme digestionMethods0.000description4
- 230000012010growthEffects0.000description4
- 238000011534incubationMethods0.000description4
- 239000013642negative controlSubstances0.000description4
- 229910052757nitrogenInorganic materials0.000description4
- 239000002773nucleotideSubstances0.000description4
- 244000052769pathogenSpecies0.000description4
- 230000001717pathogenic effectEffects0.000description4
- 102000040650(ribonucleotides)n+mHuman genes0.000description3
- QKNYBSVHEMOAJP-UHFFFAOYSA-N2-amino-2-(hydroxymethyl)propane-1,3-diol;hydron;chlorideChemical compoundCl.OCC(N)(CO)COQKNYBSVHEMOAJP-UHFFFAOYSA-N0.000description3
- 238000009010Bradford assayMethods0.000description3
- 102000018700F-Box ProteinsHuman genes0.000description3
- 108010066805F-Box ProteinsProteins0.000description3
- 241000238631HexapodaSpecies0.000description3
- 108090000364LigasesProteins0.000description3
- 102000003960LigasesHuman genes0.000description3
- 102000044159UbiquitinHuman genes0.000description3
- 108090000848UbiquitinProteins0.000description3
- 150000001413amino acidsChemical class0.000description3
- 230000008859changeEffects0.000description3
- 239000003153chemical reaction reagentSubstances0.000description3
- 238000003776cleavage reactionMethods0.000description3
- 238000000749co-immunoprecipitationMethods0.000description3
- 230000029087digestionEffects0.000description3
- 239000012634fragmentSubstances0.000description3
- 230000002401inhibitory effectEffects0.000description3
- 238000002347injectionMethods0.000description3
- 239000007924injectionSubstances0.000description3
- 229930027917kanamycinNatural products0.000description3
- 229960000318kanamycinDrugs0.000description3
- SBUJHOSQTJFQJX-NOAMYHISSA-NkanamycinChemical compoundO[C@@H]1[C@@H](O)[C@H](O)[C@@H](CN)O[C@@H]1O[C@H]1[C@H](O)[C@@H](O[C@@H]2[C@@H]([C@@H](N)[C@H](O)[C@@H](CO)O2)O)[C@H](N)C[C@@H]1NSBUJHOSQTJFQJX-NOAMYHISSA-N0.000description3
- 229930182823kanamycin ANatural products0.000description3
- 239000000203mixtureSubstances0.000description3
- 125000003729nucleotide groupChemical group0.000description3
- 238000003753real-time PCRMethods0.000description3
- 230000004044responseEffects0.000description3
- JQXXHWHPUNPDRT-WLSIYKJHSA-NrifampicinChemical compoundO([C@](C1=O)(C)O/C=C/[C@@H]([C@H]([C@@H](OC(C)=O)[C@H](C)[C@H](O)[C@H](C)[C@@H](O)[C@@H](C)\C=C\C=C(C)/C(=O)NC=2C(O)=C3C([O-])=C4C)C)OC)C4=C1C3=C(O)C=2\C=N\N1CC[NH+](C)CC1JQXXHWHPUNPDRT-WLSIYKJHSA-N0.000description3
- 229960001225rifampicinDrugs0.000description3
- 239000012723sample bufferSubstances0.000description3
- 230000007017scissionEffects0.000description3
- 230000035945sensitivityEffects0.000description3
- 239000000243solutionSubstances0.000description3
- 239000000758substrateSubstances0.000description3
- 239000000725suspensionSubstances0.000description3
- 238000011426transformation methodMethods0.000description3
- 230000009385viral infectionEffects0.000description3
- 108091068561Aux/IAA familyProteins0.000description2
- 108020004705CodonProteins0.000description2
- 208000035240Disease ResistanceDiseases0.000description2
- YQYJSBFKSSDGFO-UHFFFAOYSA-NEpihygromycinNatural productsOC1C(O)C(C(=O)C)OC1OC(C(=C1)O)=CC=C1C=C(C)C(=O)NC1C(O)C(O)C2OCOC2C1OYQYJSBFKSSDGFO-UHFFFAOYSA-N0.000description2
- 241000588724Escherichia coliSpecies0.000description2
- LFQSCWFLJHTTHZ-UHFFFAOYSA-NEthanolChemical compoundCCOLFQSCWFLJHTTHZ-UHFFFAOYSA-N0.000description2
- 238000012408PCR amplificationMethods0.000description2
- 229940124158Protease/peptidase inhibitorDrugs0.000description2
- 238000012228RNA interference-mediated gene silencingMethods0.000description2
- 101710105008RNA-binding proteinProteins0.000description2
- 241000702247ReoviridaeSpecies0.000description2
- 241000519995Stachys sylvaticaSpecies0.000description2
- 101710172711Structural proteinProteins0.000description2
- OJOBTAOGJIWAGB-UHFFFAOYSA-NacetosyringoneChemical compoundCOC1=CC(C(C)=O)=CC(OC)=C1OOJOBTAOGJIWAGB-UHFFFAOYSA-N0.000description2
- 239000011543agarose gelSubstances0.000description2
- 229960000723ampicillinDrugs0.000description2
- AVKUERGKIZMTKX-NJBDSQKTSA-NampicillinChemical compoundC1([C@@H](N)C(=O)N[C@H]2[C@H]3SC([C@@H](N3C2=O)C(O)=O)(C)C)=CC=CC=C1AVKUERGKIZMTKX-NJBDSQKTSA-N0.000description2
- 239000011324beadSubstances0.000description2
- 238000005119centrifugationMethods0.000description2
- 238000007796conventional methodMethods0.000description2
- NKLPQNGYXWVELD-UHFFFAOYSA-Mcoomassie brilliant blueChemical compound[Na+].C1=CC(OCC)=CC=C1NC1=CC=C(C(=C2C=CC(C=C2)=[N+](CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=2C=CC(=CC=2)N(CC)CC=2C=C(C=CC=2)S([O-])(=O)=O)C=C1NKLPQNGYXWVELD-UHFFFAOYSA-M0.000description2
- 238000013461designMethods0.000description2
- 238000001514detection methodMethods0.000description2
- 238000011161developmentMethods0.000description2
- 230000004069differentiationEffects0.000description2
- 239000000539dimerSubstances0.000description2
- 230000000694effectsEffects0.000description2
- 239000000499gelSubstances0.000description2
- 238000001502gel electrophoresisMethods0.000description2
- 230000006698inductionEffects0.000description2
- 238000011081inoculationMethods0.000description2
- 229910001629magnesium chlorideInorganic materials0.000description2
- 239000003550markerSubstances0.000description2
- 239000000463materialSubstances0.000description2
- 230000034217membrane fusionEffects0.000description2
- 230000004048modificationEffects0.000description2
- 238000012986modificationMethods0.000description2
- 230000036961partial effectEffects0.000description2
- 239000008188pelletSubstances0.000description2
- 239000000137peptide hydrolase inhibitorSubstances0.000description2
- 239000013641positive controlSubstances0.000description2
- 230000029279positive regulation of transcription, DNA-dependentEffects0.000description2
- 230000001681protective effectEffects0.000description2
- 238000011084recoveryMethods0.000description2
- 238000003757reverse transcription PCRMethods0.000description2
- 239000011780sodium chlorideSubstances0.000description2
- 239000002689soilSubstances0.000description2
- 239000008223sterile waterSubstances0.000description2
- 238000013518transcriptionMethods0.000description2
- 230000035897transcriptionEffects0.000description2
- 230000002103transcriptional effectEffects0.000description2
- OWEGMIWEEQEYGQ-UHFFFAOYSA-N100676-05-9Natural productsOC1C(O)C(O)C(CO)OC1OCC1C(O)C(O)C(O)C(OC2C(OC(O)C(O)C2O)CO)O1OWEGMIWEEQEYGQ-UHFFFAOYSA-N0.000description1
- 108091061173ARF familyProteins0.000description1
- 102000041188ARF familyHuman genes0.000description1
- 108010022579ATP dependent 26S proteaseProteins0.000description1
- 102000007469ActinsHuman genes0.000description1
- 108010085238ActinsProteins0.000description1
- 229920000856AmylosePolymers0.000description1
- 241000219194ArabidopsisSpecies0.000description1
- 241000894006BacteriaSpecies0.000description1
- 108091026890Coding regionProteins0.000description1
- 108700010070Codon UsageProteins0.000description1
- 102000012410DNA LigasesHuman genes0.000description1
- 108010061982DNA LigasesProteins0.000description1
- 102100031438E3 ubiquitin-protein ligase RING1Human genes0.000description1
- 108010068370GlutensProteins0.000description1
- 101000707962Homo sapiens E3 ubiquitin-protein ligase RING1Proteins0.000description1
- 102000003918Hyaluronan SynthasesHuman genes0.000description1
- 108090000320Hyaluronan SynthasesProteins0.000description1
- 229910009891LiAcInorganic materials0.000description1
- GUBGYTABKSRVRQ-PICCSMPSSA-NMaltoseNatural productsO[C@@H]1[C@@H](O)[C@H](O)[C@@H](CO)O[C@@H]1O[C@@H]1[C@@H](CO)OC(O)[C@H](O)[C@H]1OGUBGYTABKSRVRQ-PICCSMPSSA-N0.000description1
- 101710082493Minor outer capsid protein P9Proteins0.000description1
- 101710159910Movement proteinProteins0.000description1
- 208000009869Neu-Laxova syndromeDiseases0.000description1
- 241000207746Nicotiana benthamianaSpecies0.000description1
- 108700005873Nicotiana glutinosa NProteins0.000description1
- 101710144117Non-structural protein 4Proteins0.000description1
- 108010077850Nuclear Localization SignalsProteins0.000description1
- 108091028043Nucleic acid sequenceProteins0.000description1
- 108090000854OxidoreductasesProteins0.000description1
- 102000004316OxidoreductasesHuman genes0.000description1
- 241000702656PhytoreovirusSpecies0.000description1
- 244000184734Pyrus japonicaSpecies0.000description1
- 238000011529RT qPCRMethods0.000description1
- 241000702263Reovirus sp.Species0.000description1
- 208000035217Ring chromosome 1 syndromeDiseases0.000description1
- 101710180274Suppressor of RNA-mediated gene silencingProteins0.000description1
- 101710120037Toxin CcdBProteins0.000description1
- 108091023040Transcription factorProteins0.000description1
- 102000040945Transcription factorHuman genes0.000description1
- 108010083111Ubiquitin-Protein LigasesProteins0.000description1
- 102000006275Ubiquitin-Protein LigasesHuman genes0.000description1
- 241000700605VirusesSpecies0.000description1
- 125000000539amino acid groupChemical group0.000description1
- 238000004458analytical methodMethods0.000description1
- 239000003242anti bacterial agentSubstances0.000description1
- 229940088710antibiotic agentDrugs0.000description1
- 230000005540biological transmissionEffects0.000description1
- UDSAIICHUKSCKT-UHFFFAOYSA-Nbromophenol blueChemical compoundC1=C(Br)C(O)=C(Br)C=C1C1(C=2C=C(Br)C(O)=C(Br)C=2)C2=CC=CC=C2S(=O)(=O)O1UDSAIICHUKSCKT-UHFFFAOYSA-N0.000description1
- 230000024245cell differentiationEffects0.000description1
- 230000032823cell divisionEffects0.000description1
- 210000000170cell membraneAnatomy0.000description1
- 230000010307cell transformationEffects0.000description1
- 238000010367cloningMethods0.000description1
- 238000003501co-cultureMethods0.000description1
- 108091036078conserved sequenceProteins0.000description1
- 230000018109developmental processEffects0.000description1
- 210000002249digestive systemAnatomy0.000description1
- 241001493065dsRNA virusesSpecies0.000description1
- 238000004520electroporationMethods0.000description1
- 230000013020embryo developmentEffects0.000description1
- 210000002257embryonic structureAnatomy0.000description1
- 230000009483enzymatic pathwayEffects0.000description1
- 238000001400expression cloningMethods0.000description1
- 239000011536extraction bufferSubstances0.000description1
- 239000012530fluidSubstances0.000description1
- IXORZMNAPKEEDV-UHFFFAOYSA-Ngibberellic acid GA3Natural productsOC(=O)C1C2(C3)CC(=C)C3(O)CCC2C2(C=CC3O)C1C3(C)C(=O)O2IXORZMNAPKEEDV-UHFFFAOYSA-N0.000description1
- 239000003292glueSubstances0.000description1
- 235000021312glutenNutrition0.000description1
- 229930182470glycosideNatural products0.000description1
- 150000002338glycosidesChemical class0.000description1
- 239000003617indole-3-acetic acidSubstances0.000description1
- 239000003112inhibitorSubstances0.000description1
- BPHPUYQFMNQIOC-NXRLNHOXSA-Nisopropyl beta-D-thiogalactopyranosideChemical compoundCC(C)S[C@@H]1O[C@H](CO)[C@H](O)[C@H](O)[C@H]1OBPHPUYQFMNQIOC-NXRLNHOXSA-N0.000description1
- 230000020673lateral root developmentEffects0.000description1
- 230000003902lesionEffects0.000description1
- 239000002502liposomeSubstances0.000description1
- 210000001161mammalian embryoAnatomy0.000description1
- 230000001404mediated effectEffects0.000description1
- 239000012528membraneSubstances0.000description1
- 229960002523mercuric chlorideDrugs0.000description1
- LWJROJCJINYWOX-UHFFFAOYSA-Lmercury dichlorideChemical compoundCl[Hg]ClLWJROJCJINYWOX-UHFFFAOYSA-L0.000description1
- 238000002156mixingMethods0.000description1
- 238000007500overflow downdraw methodMethods0.000description1
- 230000037361pathwayEffects0.000description1
- 230000008121plant developmentEffects0.000description1
- 230000008635plant growthEffects0.000description1
- 238000002264polyacrylamide gel electrophoresisMethods0.000description1
- 238000001556precipitationMethods0.000description1
- 230000008569processEffects0.000description1
- 230000009465prokaryotic expressionEffects0.000description1
- 238000000751protein extractionMethods0.000description1
- 230000004850protein–protein interactionEffects0.000description1
- 230000009467reductionEffects0.000description1
- 239000011347resinSubstances0.000description1
- 229920005989resinPolymers0.000description1
- 239000012882rooting mediumSubstances0.000description1
- 230000035939shockEffects0.000description1
- 230000011664signalingEffects0.000description1
- 239000012879subculture mediumSubstances0.000description1
- 239000008399tap waterSubstances0.000description1
- 235000020679tap waterNutrition0.000description1
- 230000006032tissue transformationEffects0.000description1
- 108091006107transcriptional repressorsProteins0.000description1
- 230000009466transformationEffects0.000description1
- 230000001131transforming effectEffects0.000description1
- 230000001052transient effectEffects0.000description1
- 230000034512ubiquitinationEffects0.000description1
- 238000010798ubiquitinationMethods0.000description1
- 210000003462veinAnatomy0.000description1
- 210000005253yeast cellAnatomy0.000description1
- 238000001086yeast two-hybrid systemMethods0.000description1
- DGVVWUTYPXICAM-UHFFFAOYSA-Nβ‐MercaptoethanolChemical compoundOCCSDGVVWUTYPXICAM-UHFFFAOYSA-N0.000description1
Images






Classifications
- C—CHEMISTRY; METALLURGY
- C07—ORGANIC CHEMISTRY
- C07K—PEPTIDES
- C07K14/00—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof
- C07K14/415—Peptides having more than 20 amino acids; Gastrins; Somatostatins; Melanotropins; Derivatives thereof from plants
- C—CHEMISTRY; METALLURGY
- C12—BIOCHEMISTRY; BEER; SPIRITS; WINE; VINEGAR; MICROBIOLOGY; ENZYMOLOGY; MUTATION OR GENETIC ENGINEERING
- C12N—MICROORGANISMS OR ENZYMES; COMPOSITIONS THEREOF; PROPAGATING, PRESERVING, OR MAINTAINING MICROORGANISMS; MUTATION OR GENETIC ENGINEERING; CULTURE MEDIA
- C12N15/00—Mutation or genetic engineering; DNA or RNA concerning genetic engineering, vectors, e.g. plasmids, or their isolation, preparation or purification; Use of hosts therefor
- C12N15/09—Recombinant DNA-technology
- C12N15/63—Introduction of foreign genetic material using vectors; Vectors; Use of hosts therefor; Regulation of expression
- C12N15/79—Vectors or expression systems specially adapted for eukaryotic hosts
- C12N15/82—Vectors or expression systems specially adapted for eukaryotic hosts for plant cells, e.g. plant artificial chromosomes (PACs)
- C12N15/8241—Phenotypically and genetically modified plants via recombinant DNA technology
- C12N15/8261—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield
- C12N15/8271—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance
- C12N15/8279—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for biotic stress resistance, pathogen resistance, disease resistance
- C12N15/8283—Phenotypically and genetically modified plants via recombinant DNA technology with agronomic (input) traits, e.g. crop yield for stress resistance, e.g. heavy metal resistance for biotic stress resistance, pathogen resistance, disease resistance for virus resistance
Landscapes
- Health & Medical Sciences (AREA)
- Life Sciences & Earth Sciences (AREA)
- Genetics & Genomics (AREA)
- Chemical & Material Sciences (AREA)
- Organic Chemistry (AREA)
- Engineering & Computer Science (AREA)
- Molecular Biology (AREA)
- Biophysics (AREA)
- General Health & Medical Sciences (AREA)
- Zoology (AREA)
- Biochemistry (AREA)
- Wood Science & Technology (AREA)
- Bioinformatics & Cheminformatics (AREA)
- General Engineering & Computer Science (AREA)
- Biotechnology (AREA)
- Biomedical Technology (AREA)
- Botany (AREA)
- Physics & Mathematics (AREA)
- Cell Biology (AREA)
- Virology (AREA)
- Plant Pathology (AREA)
- Gastroenterology & Hepatology (AREA)
- Microbiology (AREA)
- Proteomics, Peptides & Aminoacids (AREA)
- Medicinal Chemistry (AREA)
- Breeding Of Plants And Reproduction By Means Of Culturing (AREA)
- Micro-Organisms Or Cultivation Processes Thereof (AREA)
Abstract
Translated fromChineseDescription
Translated fromChinese技术领域technical field
本发明涉及生物技术领域,尤其涉及降低IAA10蛋白及其编码基因表达在提高植物对水稻矮缩病毒抗性中的应用。The invention relates to the field of biotechnology, in particular to the application of reducing the expression of IAA10 protein and its encoded gene in improving the resistance of plants to rice dwarf virus.
背景技术Background technique
生长素在植物生长发育的很多方面发挥作用,如细胞的分裂和分化、胚的发育、顶端优势的形成、茎的伸长、侧根的发育等。生长素能够在很短时间内改变其所调控的基因的表达。有3类基因能够在施加生长素后被迅速诱导表达,它们分别为:Aux/IAA(AUXIN/INDOLE-3-ACETIC ACID)家族、GH3(GRETCHENHAGEN-3)家族和SAUR(small auxin up RNA)家族。Aux/IAA家族基因编码一类半衰期很短的转录抑制子。在生长素浓度很低时,它们的转录水平也很低,在生长素水平升高后,它们被迅速诱导表达,因此这类基因在生长素的快速瞬时反应中起关键作用。Auxin plays a role in many aspects of plant growth and development, such as cell division and differentiation, embryo development, formation of apical dominance, stem elongation, and lateral root development. Auxin can change the expression of the genes it regulates in a very short time. There are three types of genes that can be rapidly induced to express after the application of auxin, they are: Aux/IAA (AUXIN/INDOLE-3-ACETIC ACID) family, GH3 (GRETCHENHAGEN-3) family and SAUR (small auxin up RNA) family . Aux/IAA family genes encode a class of transcriptional repressors with short half-lives. Their transcription levels are also low when auxin concentrations are low, and their expression is rapidly induced after auxin levels rise, so such genes play a key role in the rapid transient response to auxin.
Aux/IAA家族蛋白很庞大,在水稻中已经发现的该家族成员至少有31个,拟南芥中则有29个。虽然IAA家族蛋白的氨基酸序列差异很大,但是它们的结构却具有很高的相似性,例如它们都具有核定位信号,大部分成员都具有4个保守的结构域(Domain I、DomainII、Domain III和Domain IV)。Doman I含有保守的LxLxL序列,具有转录抑制活性,它不仅能够抑制与之融合的蛋白的转录激活活性,而且能够抑制与它相互作用的蛋白的转录激活活性。Domain II包含一段高度保守的序列,能够被该蛋白泛素化过程中的E3识别,与IAA蛋白的降解相关。Domain III和Domain IV是介导蛋白-蛋白相互作用的区域,能够使同一种IAA蛋白或者不同种IAA蛋白形成二聚体,还能使IAA蛋白与ARF(auxin response factor)蛋白形成二聚体。因此IAA家族蛋白是一类半衰期很短的具有转录抑制活性的蛋白因子。The Aux/IAA family of proteins is very large. At least 31 members of this family have been found in rice and 29 in Arabidopsis. Although the amino acid sequences of IAA family proteins are very different, their structures share high similarity, for example, they all have nuclear localization signals, and most members have 4 conserved domains (Domain I, Domain II, Domain III). and Domain IV). Doman I contains a conserved LxLxL sequence and has transcriptional inhibitory activity. It can not only inhibit the transcriptional activation activity of the protein fused with it, but also inhibit the transcriptional activation activity of the protein that interacts with it. Domain II contains a highly conserved sequence that is recognized by E3 during ubiquitination of this protein and is associated with the degradation of IAA protein. Domain III and Domain IV are regions that mediate protein-protein interactions, which can make the same IAA protein or different IAA proteins form dimers, and can also make IAA proteins form dimers with ARF (auxin response factor) proteins. Therefore, IAA family proteins are a class of protein factors with short half-life and transcriptional inhibitory activity.
IAA蛋白是auxin通路中非常关键的因子。通常情况下,IAA蛋白与转录因子ARF家族成员相互作用,形成异源二聚体,抑制ARF调控的下游基因的表达。在生长素水平升高的情况下,IAA蛋白和F-box Transport Inhibitor Response/Auxin Signaling F-box(TIR1/AFB)作为auxin的共同受体,它们之间的相互作用增强。TIR1是泛素E3连接酶SCFTIR1复合体的一部分,其中TIR1是复合体的F-box,负责结合底物,然后与复合体中的其他成分配合,将泛素ubiquitin加到底物上,加上泛素串的底物可以被26S蛋白酶体识别、降解。Auxin像胶水一样,将TIR1和Aux/IAA蛋白紧密地结合在一起,使得Aux/IAA蛋白被SCFTIR1复合体泛素化,通过26S酶体途径降解,从而解除了其对ARF的抑制作用,启动一系列与生长发育及抗病相关的基因的表达。IAA protein is a very critical factor in the auxin pathway. Normally, IAA proteins interact with members of the ARF family of transcription factors to form heterodimers that inhibit the expression of ARF-regulated downstream genes. In the presence of elevated auxin levels, the interaction between IAA protein and F-box Transport Inhibitor Response/Auxin Signaling F-box (TIR1/AFB) as auxin co-receptors was enhanced. TIR1 is part of the ubiquitin E3 ligase SCFTIR1 complex, in which TIR1 is the F-box of the complex, responsible for binding the substrate, and then cooperates with other components in the complex to add ubiquitin to the substrate, plus The substrates of ubiquitin strings can be recognized and degraded by the 26S proteasome. Like glue, Auxin tightly binds TIR1 and Aux/IAA proteins together, so that Aux/IAA proteins are ubiquitinated by the SCFTIR1 complex and degraded through the 26S enzymatic pathway, thereby releasing its inhibitory effect on ARF and starting the Expression of a series of genes related to growth, development and disease resistance.
水稻矮缩病毒(Rice Dwarf Virus,RDV)是呼肠孤病毒科(Reoviridae)植物呼肠孤病毒属(Phytoreovirus)的成员。RDV是引起水稻矮缩病的病原,可以感染水稻,造成水稻严重减产。RDV的传播需借助于昆虫介体叶蝉。RDV为双链RNA病毒,其基因组为十二条双链RNA。根据这十二条双链RNA在聚丙烯酰胺凝胶电泳中的迁移率由慢到快,分别命名为S1到S12。RDV基因组编码至少七种结构蛋白,包括P1、P2、P3、P5、P7、P8、P9;五种非结构蛋白,包括Pns4、Pns6、Pns10、Pns11、Pns12。Rice dwarf virus (Rice Dwarf Virus, RDV) is a member of Reoviridae (Reoviridae) plant Reovirus (Phytoreovirus). RDV is the pathogen that causes rice dwarf disease, which can infect rice and cause severe rice yield reduction. The transmission of RDV relies on the insect vector leafhopper. RDV is a double-stranded RNA virus with twelve double-stranded RNAs in its genome. According to the mobility of these twelve double-stranded RNAs in polyacrylamide gel electrophoresis from slow to fast, they were named S1 to S12 respectively. The RDV genome encodes at least seven structural proteins, including P1, P2, P3, P5, P7, P8, and P9; and five non-structural proteins, including Pns4, Pns6, Pns10, Pns11, and Pns12.
P2是RDV的外壳蛋白,由S2编码,它能够与水稻内根-贝壳杉烯氧化酶及其类似蛋白相互作用,引起水稻赤霉素含量的降低,是造成水稻矮缩症状的原因之一(Zhu et al.,2005.The Rice Dwarf Virus P2Proteininteracts with ent-Kaurene Oxidases invivo,leading to reduced biosynthesis of Gibberellins andrice dwarfsymptoms.Plant Physiology.139:1935-1945.)。另外,P2在RDV的昆虫介体叶蝉细胞中,能够引起细胞膜的融合,说明该蛋白在RDV侵染叶蝉过程中发挥作用(Zhou et al.,2007,TheP2capsidprotein of the nonenveloped rice dwarf phytoreovirus induces membranefusion in insect host cells.Proceedings of the National Academy of Sciencesof USA.104(49):19547-19552.)。P2 is the coat protein of RDV, encoded by S2, which can interact with rice root-kaurene oxidase and its similar proteins to reduce the content of gibberellin in rice, which is one of the reasons for rice dwarf symptoms ( Zhu et al., 2005. The Rice Dwarf Virus P2Proteininteracts with ent-Kaurene Oxidases invivo, leading to reduced biosynthesis of Gibberellins andrice dwarfsymptoms.Plant Physiology.139:1935-1945.). In addition, P2 can induce cell membrane fusion in the insect mediator leafhopper cells of RDV, indicating that this protein plays a role in the process of RDV infection of leafhoppers (Zhou et al., 2007, TheP2capsidprotein of the nonenveloped rice dwarf phytoreovirus induces membranefusion in insect host cells. Proceedings of the National Academy of Sciences of USA. 104(49):19547-19552.).
发明内容SUMMARY OF THE INVENTION
本发明的一个目的是提供一种用于调控植物抗水稻矮缩病(或提高植物对水稻矮缩病毒抗性)的蛋白IAA10,所述蛋白是:One object of the present invention is to provide a protein IAA10 for regulating plant resistance to rice dwarf disease (or improving plant resistance to rice dwarf virus), the protein is:
(A)具有序列表中的序列1所示的氨基酸序列的蛋白;(A) a protein having the amino acid sequence shown in SEQ ID NO: 1 in the sequence listing;
(B)包括与序列1所示的氨基酸序列具有95%以上,优选98%以上的同一性的氨基酸序列且具有对水稻矮缩病毒抗性的蛋白。(B) A protein comprising an amino acid sequence that is 95% or more, preferably 98% or more identical to the amino acid sequence shown in SEQ ID NO: 1, and has resistance to rice dwarf virus.
上述(B)的蛋白可以通过置换、取代、附加或缺失(A)的序列中一个或多个氨基酸来获得。The protein of (B) above can be obtained by substituting, substituting, adding or deleting one or more amino acids in the sequence of (A).
本发明的另一个目的是提供用于调控植物抗水稻矮缩病(或提高植物对水稻矮缩病毒抗性)的编码蛋白IAA10的基因,所述基因是:Another object of the present invention is to provide a gene encoding protein IAA10 for regulating plant resistance to rice dwarf disease (or improving plant resistance to rice dwarf virus), the gene is:
(A)具有序列表中的序列2所示的碱基序列的DNA分子;(A) a DNA molecule having the base sequence shown in SEQ ID NO: 2 in the sequence listing;
(B)编码具有序列表中的序列1所示的氨基酸序列的蛋白的DNA分子;(B) a DNA molecule encoding a protein having the amino acid sequence shown in SEQ ID NO: 1 in the Sequence Listing;
(C)编码蛋白的DNA分子,所述蛋白为包括与序列1所示的氨基酸序列具有95%以上,优选98%以上的同一性的氨基酸序列且具有对水稻矮缩病毒抗性的蛋白。(C) A DNA molecule encoding a protein comprising an amino acid sequence that is 95% or more, preferably 98% or more identical to the amino acid sequence shown in SEQ ID NO: 1, and has resistance to rice dwarf virus.
本发明的又一个目的是提供上述基因的RNAi(RNA干扰)载体。该RNAi载体例如含有反向互补形式(发夹结构)的上述编码蛋白IAA10的基因。Yet another object of the present invention is to provide RNAi (RNA interference) vectors of the above-mentioned genes. The RNAi vector contains, for example, the above-mentioned gene encoding the protein IAA10 in a reverse complementary form (hairpin structure).
所述载体可以使用现有的表达载体。现有的植物表达载体包括但不局限于pCambia2300、pCambia1300、pCambia1301、pCambia3301、pWM101等。具体重组载体是本申请实施例中制备的pCambia2300-actin-IAA10RNAi。The vector can use an existing expression vector. Existing plant expression vectors include but are not limited to pCambia2300, pCambia1300, pCambia1301, pCambia3301, pWM101 and the like. The specific recombinant vector is pCambia2300-actin-IAA10RNAi prepared in the examples of this application.
本发明还提供了构建上述表达载体的方法,该方法包括:The present invention also provides a method for constructing the above-mentioned expression vector, the method comprising:
1)获得序列2所示的DNA分子;1) obtain the DNA molecule shown in
2)根据序列2所示的DNA分子设计引物,并在引物两端加上所需酶切位点和保护碱基,引物序列为IAA10-5'BamHI:5’-GTGGATCCAAATGAGAGGAGGAGTAGCTGG-3’,IAA10-3'XhoI:5’-GCCTCGAGTCAGGATCTGCCTCTTGTTG-3’;2) Design primers according to the DNA molecule shown in
3)用Invitrogen公司的TRIzol Reagent提取中花11水稻的总RNA,用该公司的SuperScript II逆转录酶进行逆转录,得到cDNA;3) The total RNA of Zhonghua 11 rice was extracted with TRIzol Reagent of Invitrogen Company, and reverse transcribed with SuperScript II reverse transcriptase of the company to obtain cDNA;
4)以逆转录所得cDNA为模板,以上述基因特异性引物IAA10-5'BamHI和IAA10-3'XhoI进行PCR反应,得到861bp的PCR产物,该PCR产物具有序列表中序列2所示的核苷酸;4) Take reverse transcription gained cDNA as template, carry out PCR reaction with above-mentioned gene-specific primers IAA10-5'BamHI and IAA10-3'XhoI, obtain the PCR product of 861bp, and this PCR product has the nucleus shown in
5)回收PCR产物后用限制性内切酶BamHI和XhoI进行酶切,回收851bp带有粘性末端的PCR产物;将载体pGADT7用限制性内切酶EcoRI和Sal I双酶切,回收7987bp载体骨架;将上述851bp带有粘性末端的PCR产物与7987bp载体骨架用T4连接酶连接,转化大肠杆菌菌株DH5α,得到转化子,提取转化子的质粒送去测序,该质粒为将序列表中序列2所示的基因IAA10插入载体pGADT7的BamHI和XhoI酶切位点间得到的载体,将该质粒命名为pGAD-IAA10,即为重组载体;5) After the recovery of the PCR product, the restriction enzymes BamHI and XhoI were used for digestion, and the 851bp PCR product with sticky ends was recovered; the vector pGADT7 was double-digested with the restriction enzymes EcoRI and Sal I, and the 7987bp vector backbone was recovered. The above-mentioned 851bp PCR product with sticky ends is connected with the 7987bp carrier backbone with T4 ligase, transforms Escherichia coli strain DH5α, obtains the transformant, and the plasmid of the extract transformant is sent to ordering, and this plasmid is the
6)以pGAD-IAA10为模版,IAA10-5'SpeI(序列为:5’-GTACTAGTATGAGAGGAGGAGTAGCTGG-3’和IAA10-3'BglII(序列为:5’-GCAGATCTTCAGGATCTGCCTCTTGTTG-3’)进行PCR扩增,得到859bp的PCR产物,将该PCR产物用SpeI和BglII双酶切,连入到用XbalI和BamHI(和SpeI和BglII为同尾酶)酶切过的中间载体pUCCRNAi上,经过测序,重组质粒pUCC-IAA10为将IAA10插入pUCCRNAi质粒的XbalI和BamHI酶切位点间得到的载体,再将用SpeI和BglII双酶切的PCR产物连入到用SpeI和BglII双酶切过pUCC-IAA10质粒中,得到pUCC-IAA10RNAi载体,经过测序,重组质粒pUCC-IAA10RNAi为将IAA10以反向互补形式插入pUCCRNAi质粒的XbalI和BamHI酶切位点和SpeI和BglII酶切位点间得到的载体;6) Using pGAD-IAA10 as a template, IAA10-5'SpeI (sequence: 5'-GTACTAGTATGAGAGGAGGAGTAGCTGG-3' and IAA10-3'BglII (sequence: 5'-GCAGATCTTCAGGATCTGCCTCTTGTTG-3') were amplified by PCR to obtain 859bp The PCR product was double digested with SpeI and BglII, and connected to the intermediate vector pUCCRNAi digested with XbalI and BamHI (with SpeI and BglII being homozygous enzymes), and after sequencing, the recombinant plasmid pUCC-IAA10 In order to insert IAA10 into the vector obtained between the XbalI and BamHI restriction sites of pUCCRNAi plasmid, the PCR product double digested with SpeI and BglII was connected into the pUCC-IAA10 plasmid double digested with SpeI and BglII to obtain pUCC -IAA10RNAi vector, after sequencing, the recombinant plasmid pUCC-IAA10RNAi is a vector obtained by inserting IAA10 into the XbalI and BamHI restriction sites and the SpeI and BglII restriction sites of the pUCCRNAi plasmid in the form of reverse complement;
7)将pUCC-IAA10RNAi载体用PstI单酶切,琼脂糖凝胶回收得到1939bp酶切产物,将该产物连接入用PstI单酶切得到的10379bp的pCambia2300-Actin1-ocs载体骨架上,得到重组载体pCambia2300-actin-IAA10RNAi。7) The pUCC-IAA10RNAi vector was digested with PstI single enzyme, and the agarose gel was recovered to obtain the 1939bp digested product, and the product was connected to the 10379bp pCambia2300-Actin1-ocs carrier backbone obtained by single enzyme digestion with PstI to obtain a recombinant vector pCambia2300-actin-IAA10RNAi.
本发明的再一个目的提供了上述蛋白IAA10或其编码基因或所述编码基因的RNAi(RNA干扰)载体在提高植物对水稻矮缩病毒抗性中(或在调控植物抗水稻矮缩病中)的应用。Another object of the present invention provides the above-mentioned protein IAA10 or its encoding gene or the RNAi (RNA interference) vector of said encoding gene in improving plant resistance to rice dwarf virus (or in regulating plant resistance to rice dwarf disease) Applications.
上述应用中,所述水稻矮缩病的病原为水稻矮缩病毒(Rice dwarf virus)。In the above application, the pathogen of the rice dwarf disease is rice dwarf virus (Rice dwarf virus).
上述应用中,所述植物优选为单子叶植物或双子叶植物,上述单子叶植物为水稻。In the above application, the plant is preferably a monocotyledonous plant or a dicotyledonous plant, and the above-mentioned monocotyledonous plant is rice.
本发明的另一个目的是提供一种培育转基因植物的方法,所述包括如下步骤:将上述蛋白IAA10的编码基因导入目的植物中,得到转基因植物,所述转基因植物的水稻矮缩病抗性低于所述目的植物;或者将上述基因以发夹结构形式转入目的植物中,得到其RNAi的转基因植物,所述转基因植物的水稻矮缩病抗性高于所述目的植物。Another object of the present invention is to provide a method for cultivating transgenic plants, comprising the steps of: introducing the encoding gene of the above-mentioned protein IAA10 into a target plant to obtain a transgenic plant, the transgenic plant having low resistance to rice dwarf disease into the target plant; or the above gene is transformed into the target plant in the form of a hairpin structure to obtain a transgenic plant with RNAi thereof, and the resistance of the transgenic plant to rice dwarf disease is higher than that of the target plant.
上述方法中,所述蛋白IAA10的编码基因的核苷酸序列为序列表中的序列2,或者为编码具有序列表中的序列1所示的氨基酸序列的蛋白的DNA分子,或者编码蛋白的DNA分子,所述蛋白为包括与序列1所示的氨基酸序列具有95%以上,优选98%以上的同一性的氨基酸序列且具有对水稻矮缩病毒抗性的蛋白。In the above method, the nucleotide sequence of the encoding gene of the protein IAA10 is the
上述方法中,所述水稻矮缩病的病原菌为水稻矮缩病毒(Rice Dwarf Virus)。In the above method, the pathogen of rice dwarf disease is Rice Dwarf Virus.
上述方法中,所述蛋白IAA10的编码基因通过重组载体导入目的植物。In the above method, the gene encoding the protein IAA10 is introduced into the target plant through a recombinant vector.
所述重组载体为将所述蛋白IAA10的编码基因插入表达载体中得到的重组载体。例如通过以上所述方法制备的重组载体。上述方法中,所述植物为单子叶植物或双子叶植物,上述单子叶植物为水稻。The recombinant vector is obtained by inserting the encoding gene of the protein IAA10 into an expression vector. For example, recombinant vectors prepared by the methods described above. In the above method, the plant is a monocotyledonous plant or a dicotyledonous plant, and the above-mentioned monocotyledonous plant is rice.
所述转基因植物的水稻矮缩病抗性高于所述目的植物具体体现在接RDV后,所述转基因植物的感染的RDV病毒的植株数目少于所述目的植物;可通过鉴定是否含有RDV病毒的目的基因(S2的部分片段)或者植株的感病特征来确定感染的RDV病毒的植株。The resistance to rice dwarf disease of the transgenic plant is higher than that of the target plant, which is specifically reflected in that after receiving RDV, the number of RDV virus-infected plants in the transgenic plant is less than that of the target plant; it can be determined by identifying whether it contains RDV virus The target gene (partial fragment of S2) or the susceptibility characteristics of the plant to determine the RDV virus-infected plant.
上述蛋白IAA10在作为蛋白-泛素连接酶中的应用也是本发明保护的范围。The application of the above-mentioned protein IAA10 as a protein-ubiquitin ligase is also within the protection scope of the present invention.
其中植物表达载体包括但不局限于pCambia2300、pCambia1300、pCambia1301、pCambia3301、pWM101等;水稻品种优选为对RDV敏感的品种如中花11、秀水11、日本晴、武3等;水稻组织或细胞的转化方法可选自农杆菌介导法、基因枪法、电击法、花粉管导入法、脂质体融合法以及其他任意可将质粒导入的方法。The plant expression vectors include but are not limited to pCambia2300, pCambia1300, pCambia1301, pCambia3301, pWM101, etc.; rice varieties are preferably RDV-sensitive varieties such as
本领域的技术人员懂得,通过置换、取代、附加或缺失蛋白序列中的一个或多个氨基酸,可对其进行修饰而不改变其功能。因此,本发明应该被理解为包括了对序列1所示的(IAA10)蛋白进行的上述修饰。Those skilled in the art understand that by substituting, substituting, adding, or deleting one or more amino acids in a protein sequence, it can be modified without altering its function. Therefore, the present invention should be understood to include the above modifications to the (IAA10) protein shown in SEQ ID NO: 1.
IAA10基因的碱基序列不局限于序列表中序列2所示,还包括具有将IAA10及带有上述修饰的IAA10蛋白中各氨基酸残基对应的任一密码子进行选择并组合的DNA序列。该密码子的选择可以按照常规方法,也可以参考宿主的密码子偏好型。The base sequence of the IAA10 gene is not limited to that shown in SEQ ID NO: 2 in the Sequence Listing, but also includes a DNA sequence that selects and combines any codon corresponding to each amino acid residue in IAA10 and the IAA10 protein with the above modifications. The selection of the codons can be carried out in accordance with conventional methods, and can also refer to the codon preference type of the host.
本发明的实验证明,本发明发现RDV外壳蛋白P2能够与IAA10相互作用,提高IAA10的稳定性,促进病毒侵染,而将IAA10表达量通过RNAi的方式降低的水稻,对RDV的抗性增强。The experiment of the present invention proves that the present invention finds that the RDV coat protein P2 can interact with IAA10, improve the stability of IAA10, and promote virus infection, and rice whose IAA10 expression is reduced by RNAi has enhanced resistance to RDV.
附图说明Description of drawings
图1为P2和IAA10蛋白在酵母中相互作用的结果;Figure 1 shows the results of the interaction between P2 and IAA10 proteins in yeast;
图2为免疫共沉淀实验验证P2和IAA10相互作用的结果;Figure 2 shows the results of co-immunoprecipitation experiments to verify the interaction between P2 and IAA10;
图3为体外降解实验证明RDV的存在使得IAA10稳定性增强;Figure 3 shows that the in vitro degradation experiment proves that the presence of RDV enhances the stability of IAA10;
图4为体外降解实验证明P2的存在使得IAA10稳定性增强;Figure 4 shows that the in vitro degradation experiment proves that the presence of P2 enhances the stability of IAA10;
图5为转IAA10水稻的Western鉴定;Fig. 5 is the Western identification of trans-IAA10 rice;
图6为转IAA10RNAi水稻的Real-time鉴定;Figure 6 is the Real-time identification of trans-IAA10RNAi rice;
图7为转IAA10和IAA10RNAi水稻和野生型水稻在RDV侵染后的RDV病毒相关基因的RT-PCR检测;Figure 7 is the RT-PCR detection of RDV virus-related genes after RDV infection in IAA10 and IAA10 RNAi rice and wild-type rice;
图8为感染RDV的不同水稻株系症状图。Figure 8 is a symptom map of different rice lines infected with RDV.
具体实施方式Detailed ways
下述实施例中所使用的实验方法如无特殊说明,均为常规方法。The experimental methods used in the following examples are conventional methods unless otherwise specified.
下述实施例中所用的材料、试剂等,如无特殊说明,均可从商业途径得到。The materials, reagents, etc. used in the following examples can be obtained from commercial sources unless otherwise specified.
以下将用实施例对本发明进行详细说明,但这些实施例并非对本发明的限制。The present invention will be described in detail below with examples, but these examples are not intended to limit the present invention.
实施例1、IAA10蛋白及其编码基因的获得Example 1. Acquisition of IAA10 protein and its encoding gene
一、IAA10蛋白及其编码基因的获得1. Acquisition of IAA10 protein and its encoding gene
水稻文库的构建所用试剂盒为HybriZAP-2.1Two-HybridPredigested Vector/GigapackCloning Kit(Stratagene),文库构建及筛选方法根据说明书进行。所用诱饵克隆pGBK-S2与专利号ZL200510114386.X所公布克隆相同。所用酵母菌株为AH109。筛选得到的阳性克隆经测序和序列分析,确定其编码的蛋白片段具有序列表中序列1所示的氨基酸序列的91-280个氨基酸,将该蛋白全长(即序列1对应蛋白)命名为IAA10,其编码基因为序列表中序列2所示的核苷酸,将其命名为IAA10。The kit used for the construction of the rice library was HybriZAP-2.1Two-HybridPredigested Vector/GigapackCloning Kit (Stratagene), and the library construction and screening methods were carried out according to the instructions. The used bait clone pGBK-S2 is the same as the clone published in Patent No. ZL200510114386.X. The yeast strain used was AH109. The positive clone obtained by screening was sequenced and sequenced, and it was confirmed that the encoded protein fragment had 91-280 amino acids of the amino acid sequence shown in SEQ ID NO: 1 in the sequence table, and the full length of the protein (that is, the corresponding protein of SEQ ID NO: 1) was named IAA10 , whose coding gene is the nucleotide shown in SEQ ID NO: 2 in the sequence listing, and is named IAA10.
二、IAA10全长序列的克隆及含有该片段的重组载体的获得2. Cloning of the full-length sequence of IAA10 and the acquisition of the recombinant vector containing the fragment
根据序列2设计引物,并在引物两端加上所需酶切位点和保护碱基,引物序列为IAA10-5'BamHI:5’-GTGGATCCAAATGAGAGGAGGAGTAGCTGG-3’,IAA10-3'XhoI:5’-GCCTCGAGTCAGGATCTGCCTCTTGTTG-3’。Design primers according to
依照说明书,用Invitrogen公司的TRIzol Reagent提取中花11水稻(Oryzasativa L.japonica cv.Zhonghua 11,许昱等《“中花11”水稻谷蛋白Gt1基因克隆及蜡质基因启动子引导Gt1基因表达载体的构建》,上海师范大学学报(自然科学版),2010年4月第39卷第2期,204页,公众可从北京大学获得)的总RNA,用该公司的SuperScript II逆转录酶进行逆转录,得到cDNA。逆转录所用引物为16个核苷酸的Oligod(T)引物。According to the instructions, use Invitrogen's TRIzol Reagent to extract
以逆转录所得cDNA为模板,以上述基因特异性引物IAA10-5'BamHI和IAA10-3'XhoI进行PCR(Polymerase Chain Reaction)反应,得到861bp的PCR产物,该PCR产物具有序列表中序列2所示的核苷酸。Take the cDNA obtained by reverse transcription as a template, carry out PCR (Polymerase Chain Reaction) reaction with above-mentioned gene-specific primers IAA10-5'BamHI and IAA10-3'XhoI, and obtain the PCR product of 861bp, and this PCR product has
回收PCR产物后用限制性内切酶BamHI和XhoI进行酶切,回收851bp带有粘性末端的PCR产物;将载体pGADT7(Clontech公司,产品目录号:630442)用限制性内切酶EcoRI和Sal I双酶切,回收7987bp载体骨架;将上述851bp带有粘性末端的PCR产物与7987bp载体骨架用T4连接酶连接,转化大肠杆菌菌株DH5α,得到转化子。提取转化子的质粒送去测序,该质粒为将序列表中序列2所示的基因IAA10插入载体pGADT7的BamHI和XhoI酶切位点间得到的载体,将该质粒命名为pGAD-IAA10,即为重组载体。After the recovery of the PCR product, the restriction enzymes BamHI and XhoI were used for digestion, and the 851bp PCR product with sticky ends was recovered; the vector pGADT7 (Clontech Company, product catalog number: 630442) was digested with the restriction enzymes EcoRI and Sal I Double-enzyme digestion was used to recover the 7987bp vector backbone; the above-mentioned 851bp PCR product with sticky ends was ligated with the7987bp vector backbone with T4 ligase, and transformed into Escherichia coli strain DH5α to obtain a transformant. The plasmid of the transformant was extracted and sent for sequencing. The plasmid was a vector obtained by inserting the gene IAA10 shown in
实施例2、体外证明IAA10蛋白与RDV P2的相互作用Example 2. In vitro demonstration of the interaction between IAA10 protein and RDV P2
一、酵母双杂交实验证明IAA10与RDV P2的相互作用1. Yeast two-hybrid experiments demonstrate the interaction between IAA10 and RDV P2
将实施例1得到的重组载体pGAD-IAA10与pGBK-S2(S2为RDV P2蛋白的编码基因,RDV P2蛋白的编码基因导入pGBK中得到的。记载该载体的非专利文献为Zhu et al.,2005.The Rice Dwarf Virus P2Proteininteracts with ent-Kaurene Oxidases invivo,leading to reduced biosynthesis of Gibberellins andrice dwarfsymptoms.Plant Physiology.139:1935-1945;公众可从北京大学获得)共转化酵母菌株AH109(购自Clontech公司,产品目录号为630444),酵母转化方法采用公知的PEG/LiAc法;转化后的酵母细胞涂布营养缺陷型平板SD/-Trp-Leu,30℃条件下培养2天。转化子划线至SD/-Trp-Leu-His-Ade营养缺陷型平板,30℃培养4天,观察转化子的生长情况。该实验同时设负对照pGADT7(产品目录号为:630442)和pGBK-S2、pGAD-IAA10和pGBK(购自Clontech公司产品目录号为630443)。The recombinant vector pGAD-IAA10 obtained in Example 1 and pGBK-S2 (S2 is the coding gene of RDV P2 protein, and the coding gene of RDV P2 protein was introduced into pGBK and obtained. The non-patent document describing this vector is Zhu et al., 2005. The Rice Dwarf Virus P2 Protein interacts with ent-Kaurene Oxidases invivo, leading to reduced biosynthesis of Gibberellins andrice dwarfsymptoms. Plant Physiology. 139: 1935-1945; publicly available from Peking University) co-transformed yeast strain AH109 (purchased from Clontech, The product catalog number is 630444), and the yeast transformation method adopts the well-known PEG/LiAc method; the transformed yeast cells are coated on an auxotrophic plate SD/-Trp-Leu, and cultured at 30° C. for 2 days. The transformants were streaked to SD/-Trp-Leu-His-Ade auxotrophic plates, cultured at 30°C for 4 days, and the growth of the transformants was observed. Negative controls pGADT7 (catalog number: 630442) and pGBK-S2, pGAD-IAA10, and pGBK (purchased from Clontech, Inc., catalog number 630443) were also used in this experiment.
结果如图1所示(A:pGAD-SV40T和pGBK-P53(阳性对照);B:pGAD-IAA10和pGBK-S2;C:pGAD-IAA10和pGBK;D:pGAD和pGBK-S2;E:pGAD-SV40T和pGBK-Lam(阴性对照))。结果证明,IAA10全长蛋白与RDVP2能在酵母中相互作用。The results are shown in Figure 1 (A: pGAD-SV40T and pGBK-P53 (positive control); B: pGAD-IAA10 and pGBK-S2; C: pGAD-IAA10 and pGBK; D: pGAD and pGBK-S2; E: pGAD - SV40T and pGBK-Lam (negative control)). The results demonstrate that the full-length protein of IAA10 interacts with RDVP2 in yeast.
二、免疫共沉淀实验证明IAA10与RDV P2的相互作用2. Co-immunoprecipitation experiments demonstrate the interaction between IAA10 and RDV P2
1)pCambia1301-FLAGIAA10及pCambia1301-HAS2克隆的构建pCambia1301-FlagIAA10载体的构建:以pGAD-IAA10为模板,以引物FlagIAA10-5'SalI(序列为:5’-CCGTCGACATGGACTACAAGGACGACGATGACAAGATGAGAGGAGGAGTAGCTG-3’)和IAA10-3’BamHI(序列为:5’-GCGGATCCTCAGGATCTGCCTCTTGTTG-3’)进行PCR扩增,得到883bp的PCR产物。将上述PCR产物用SalI和BamHI双酶切,回收酶切产物,与经过同样酶切pRTL2质粒的3852bp的载体骨架连接,得到重组质粒pRTL2-FLAGIAA10,经过测序,重组质粒pRTL2-FLAGIAA10为将FLAG-IAA10插入pRTL2质粒的SalI和BamHI酶切位点间得到的载体。1) Construction of pCambia1301-FLAGIAA10 and pCambia1301-HAS2 clones Construction of pCambia1301-FlagIAA10 vector: using pGAD-IAA10 as a template, with primers FlagIAA10-5'SalI (sequence: 5'-CCGTCGACATGGACTACAAGGACGACGATGACAAGATGAGAGGAGGAGTAGCTG-3') and IAA10-3' BamHI (sequence: 5'-GCGGATCCTCAGGATCTGCCTCTTGTTG-3') was amplified by PCR to obtain a PCR product of 883 bp. The above-mentioned PCR product is double digested with SalI and BamHI, and the digested product is recovered, and is connected with the carrier backbone of 3852bp through the same digested pRTL2 plasmid to obtain the recombinant plasmid pRTL2-FLAGIAA10. After sequencing, the recombinant plasmid pRTL2-FLAGIAA10 is the FLAG- The vector obtained by inserting IAA10 between the SalI and BamHI restriction sites of the pRTL2 plasmid.
然后将pRTL2-FLAGIAA10用HindIII单酶切,得到1985bp的酶切产物,回收酶切产物,与用同样酶酶切的pCambia1301质粒(参见Zhu et al.,2005.The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases in vivo,leading to reducedbiosynthesis of Gibberellins andrice dwarf symptoms.Plant Physiology.139:1935-1945.公众可从北京大学获得),得到11837bp的载体骨架连接,得到重组载体pCambia1301-FlagIAA10(HindIII单酶切,得到1985bp的酶切产物为阳性)。Then pRTL2-FLAGIAA10 was digested with HindIII to obtain a 1985bp digested product, and the digested product was recovered, which was compared with the pCambia1301 plasmid digested with the same enzyme (see Zhu et al., 2005. The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases). in vivo, leading to reduced biosynthesis of Gibberellins andrice dwarf symptoms. Plant Physiology. 139: 1935-1945. Publicly available from Peking University), a 11837 bp vector backbone was obtained and the recombinant vector pCambia1301-FlagIAA10 (HindIII single digestion, obtained 1985 bp) cleavage product was positive).
pCambia1301-HAS2(记载该载体的非专利文献为:Zhu et al.,2005.The RiceDwarf Virus P2Proteininteracts with ent-Kaurene Oxidases in vivo,leading toreduced biosynthesis of Gibberellins andrice dwarf symptoms.PlantPhysiology.139:1935-1945;公众可从北京大学获得),其为将HAS2(HAP2是在P2蛋白上带了HA标签)构建入pCambia1301。pCambia1301-HAS2 (non-patent literature describing this vector is: Zhu et al., 2005. The RiceDwarf Virus P2Proteininteracts with ent-Kaurene Oxidases in vivo, leading toreduced biosynthesis of Gibberellins andrice dwarf symptoms.PlantPhysiology.139:1935-1945;Public available from Peking University), which is the construction of HAS2 (HAP2 is HA-tagged on the P2 protein) into pCambia1301.
pCambia1301-FLAGIAA10和pCambia1301-HAS2分别转化农杆菌EHA105,转化方法为公知的电击转化方法,得到EHA105/pCambia1301-FLAGIAA10和EHA105/pCambia1301-HAS2。Agrobacterium EHA105 was transformed with pCambia1301-FLAGIAA10 and pCambia1301-HAS2, respectively, and the transformation method was a known electroporation method to obtain EHA105/pCambia1301-FLAGIAA10 and EHA105/pCambia1301-HAS2.
2)HAP2和FLAGIAA10在烟草中的表达2) Expression of HAP2 and FLAGIAA10 in tobacco
分别挑取EHA105/pCambia1301-FLAGIAA10和EHA105/pCambia1301-HAS2单菌落,接入2ml LB液体培养基(含有Kanamycin 100mg/L,Rifampicin 50mg/L),28℃振荡培养16hr(hr为小时hour的简写)。1:50转接入10ml LB液体培养基(含有Kanamycin 100mg/L,Rifampicin50mg/L),继续培养至OD600约为0.8。4000rpm(rpm为每分钟转速)离心5min(min为分钟)收菌。AS buffer(10mM MES,150μM AS,10mM MgCl2)重悬,调OD600至1.0。Pick out a single colony of EHA105/pCambia1301-FLAGIAA10 and EHA105/pCambia1301-HAS2 respectively, insert into 2ml LB liquid medium (containing 100mg/L of Kanamycin, 50mg/L of Rifampicin), and culture with shaking at 28°C for 16hr (hr is the abbreviation of hour) . Transfer to 10ml LB liquid medium (containing 100mg/L of Kanamycin, 50mg/L of Rifampicin) at 1:50, and continue to culture until the OD600 is about 0.8. Centrifuge at 4000rpm (rpm is the rotation speed per minute) for 5min (min is the minute) to collect bacteria. Resuspend in AS buffer (10 mM MES, 150 μM AS, 10 mM MgCl2 ) and adjust OD600 to 1.0.
将两种菌液以1:1(体积)混合,通过压力注射方法注射烟草N.benthamiana(记载该烟草的非专利文献为Zhu et al.,2005.The Rice Dwarf Virus P2Proteininteractswith ent-Kaurene Oxidases in vivo,leading to reduced biosynthesis ofGibberellins andrice dwarf symptoms.Plant Physiology.139:1935-1945.公众可从北京大学获得)并分别单独取两种菌悬液通过压力注射方法注射烟草作为负对照。烟草继续生长3天后,将注射的叶片置于液氮中研磨成粉末。Mix the two bacterial liquids at 1:1 (volume), and inject tobacco N. benthamiana by pressure injection method (the non-patent document describing the tobacco is Zhu et al., 2005. The Rice Dwarf Virus P2Proteininteractswith ent-Kaurene Oxidases in vivo. , leading to reduced biosynthesis of Gibberellins andrice dwarf symptoms. Plant Physiology. 139: 1935-1945. The public can be obtained from Peking University) and two bacterial suspensions were taken separately and injected into tobacco by pressure injection method as a negative control. After the tobacco continued to grow for 3 days, the injected leaves were placed in liquid nitrogen and ground to a powder.
3)免疫共沉淀实验3) Co-immunoprecipitation experiment
取约500mg上述叶片粉末加入1.5ml离心管中,加入1ml IP Buffer(150mM NaCl,50mM Tris-Cl(pH7.5),0.1%NP-40,5mM DTT,每50ml加1片蛋白酶抑制剂(ProteaseInhibitor Cocktail Tablets,EDTA-free;Roche),混匀后4℃,12000rpm离心10min,将上清吸出转移至新的离心管中。再次以上面条件离心5min,将上清吸出转移至新的离心管中,防止吸到沉淀,得到烟草蛋白提取物。Take about 500 mg of the above leaf powder into a 1.5 ml centrifuge tube, add 1 ml of IP Buffer (150 mM NaCl, 50 mM Tris-Cl (pH 7.5), 0.1% NP-40, 5 mM DTT, and 1 tablet of Protease Inhibitor (Protease Inhibitor) per 50 ml. Cocktail Tablets, EDTA-free; Roche), after mixing, centrifuge at 12000rpm for 10min at 4°C, transfer the supernatant to a new centrifuge tube. Centrifuge again under the above conditions for 5min, and transfer the supernatant to a new centrifuge tube. , to prevent inhalation of precipitation to obtain tobacco protein extract.
在上述步骤中得到的烟草蛋白提取物中加入20μlprotein G beads(GEhealthcare,17-0618-06),同时加入0.2μl FLAG抗体(Sigma,F3165),4℃温育1.5hr。4℃,1000rpm离心1min,吸掉上清。用IP Buffer 洗beads3次。加入50μl 3×protein samplebuffer(125mM Tris-Cl(pH6.8),20%甘油,6%SDS,0.004%溴酚蓝,30mM DTT),95℃加热5min,吸取上清进行SDS-PAGE。20 μl protein G beads (GEhealthcare, 17-0618-06) were added to the tobacco protein extract obtained in the above step, and 0.2 μl FLAG antibody (Sigma, F3165) was added at the same time, and incubated at 4° C. for 1.5 hr. Centrifuge at 1000 rpm for 1 min at 4°C and aspirate the supernatant. Wash the
SDS-PAGE及Western Blot依照公知方法及产品说明书进行。所用抗体为anti-FLAG-HRP(sigma)和anti-HA-Peroxidase(Roche)。SDS-PAGE and Western Blot were performed according to known methods and product specifications. Antibodies used were anti-FLAG-HRP (sigma) and anti-HA-Peroxidase (Roche).
结果如图2所示,表明用FLAG抗体能够将FLAGIAA10和HAP2同时沉淀下来,说明这两种蛋白之间相互作用。The results are shown in Figure 2, indicating that FLAGIAA10 and HAP2 can be simultaneously precipitated by FLAG antibody, indicating the interaction between these two proteins.
实施例3、P2能够减弱IAA10的降解Example 3. P2 can attenuate the degradation of IAA10
一、表达克隆构建及蛋白的表达1. Expression cloning and protein expression
以pGAD-IAA10质粒为模板,用IAA10-5'SalI(序列为:5’-GTGTCGACATGAGAGGAGGAGTAGCTGG-3’和IAA10-3'HindIII(序列为:5’-GCAAGCTTTCAGGATCTGCCTCTTGTTG-3’)进行PCR扩增,得到859bp的PCR产物。将此859bp的PCR产物以SalI和HindIII双酶切后,凝胶电泳回收853bp的酶切产物;将pMAL-p2x质粒(NewEngland Biolabs,E8000S,本身表达MBP蛋白)SalI和HindIII双酶切,凝胶电泳回收6703bp的载体骨架;将853bp的酶切产物和6713bp的载体骨架以T4DNA连接酶连接,得到重组载体pMAL-p2x-IAA10,经过测序,该重组载体为将序列表中序列2所示的DNA分子插入pMAL-p2x的SalI和HindIII酶切位点间得到的载体,命名为pMAL-p2x-IAA10。Using the pGAD-IAA10 plasmid as a template, IAA10-5'SalI (sequence: 5'-GTGTCGACATGAGAGGAGGAGTAGCTGG-3' and IAA10-3'HindIII (sequence: 5'-GCAAGCTTTCAGGATCTGCCTCTTGTTG-3') were used for PCR amplification to obtain 859bp After the PCR product of this 859bp was digested with SalI and HindIII, the digested product of 853bp was recovered by gel electrophoresis; The 6703bp vector backbone was recovered by gel electrophoresis; the 853bp digested product and the 6713bp vector backbone were connected with T4 DNA ligase to obtain the recombinant vector pMAL-p2x-IAA10. After sequencing, the recombinant vector was the
将pMAL-p2x-IAA10和pMAL-p2x质粒分别转化原核表达菌株Rosetta(DE3):取0.5μl质粒,加入50μl感受态细胞中,混匀后冰上静置30min;42℃热激90sec,加入1ml新鲜LB培养基,37℃振荡培养1hr;3000rpm离心2min收集菌体,涂布LB平板(含100mg/LAmpicillin),得到Rosetta(DE3)/pMAL-p2x-IAA10(SalI和HindIII双酶切鉴定,得到853bp的酶切产物的为阳性)和Rosetta(DE3)/pMAL-p2x。Transform the pMAL-p2x-IAA10 and pMAL-p2x plasmids into the prokaryotic expression strain Rosetta (DE3) respectively: take 0.5 μl of the plasmid, add it to 50 μl of competent cells, mix well and let stand on ice for 30 min; heat shock at 42°C for 90 sec, add 1 ml Fresh LB medium was shaken at 37°C for 1 hr; centrifuged at 3000 rpm for 2 min to collect the cells, and coated with LB plates (containing 100 mg/LAmpicillin) to obtain Rosetta (DE3)/pMAL-p2x-IAA10 (SalI and HindIII double enzyme digestion identification, obtained 853bp digested product was positive) and Rosetta(DE3)/pMAL-p2x.
分别挑Rosetta(DE3)/pMAL-p2x-IAA10和Rosetta(DE3)/pMAL-p2x单菌落接于5mlLB培养基中(含100mg/L Ampicillin),37℃过夜培养。1:200转接于300ml LB培养基中(含100mg/L Ampicillin)培养至OD600约为0.6。加入IPTG至终浓度1mM,诱导表达4hr,得到Rosetta(DE3)/pMAL-p2x-IAA10菌液和Rosetta(DE3)/pMAL-p2x菌液。5000rpm离心10min收集菌体。加入5ml过柱缓冲液(20mM Tris-Cl pH7.5,100mM NaCl,0.1%NP-40,1mM DTT,0.01M PMSF)悬浮菌体,超声破碎。4℃,12000rpm,离心20min,取上清液加入到用8倍体积过柱缓冲液洗涤过的Amylose Resin Column顶,使其缓缓流过柱体。用12倍体积过柱缓冲液洗涤。用含有10mM麦芽糖的过柱缓冲液洗脱纯化的蛋白。用Bradford法测定蛋白的浓度(Bio-Rad),调节至4mg/ml。Single colonies of Rosetta(DE3)/pMAL-p2x-IAA10 and Rosetta(DE3)/pMAL-p2x were picked and plated in 5ml LB medium (containing 100mg/L Ampicillin) and cultured at 37°C overnight. 1:200 was transferred to 300ml LB medium (containing 100mg/L Ampicillin) and cultured to an OD600 of about 0.6. IPTG was added to a final concentration of 1 mM, and expression was induced for 4 hr to obtain Rosetta(DE3)/pMAL-p2x-IAA10 bacterial solution and Rosetta(DE3)/pMAL-p2x bacterial solution. Cells were collected by centrifugation at 5000 rpm for 10 min. 5 ml of column buffer (20 mM Tris-Cl pH7.5, 100 mM NaCl, 0.1% NP-40, 1 mM DTT, 0.01 M PMSF) was added to suspend the cells and sonicated. 4°C, 12000 rpm, centrifugation for 20 min, take the supernatant and add it to the top of the Amylose Resin Column washed with 8 times the volume of the column buffer to make it slowly flow through the column. Wash with 12 volumes of column buffer. Purified protein was eluted with column run buffer containing 10 mM maltose. The protein concentration was determined by Bradford method (Bio-Rad) and adjusted to 4 mg/ml.
二、MBP-IAA10在水稻总蛋白提取物中的降解实验2. Degradation experiment of MBP-IAA10 in rice total protein extract
RDV感病水稻和健康水稻在液氮中研磨成粉末,加入适量体外降解缓冲液(25mMTris-HCl,pH7.5,10mM NACl,10mM MgCl2,4mM PMSF,5mM DTT,10mM ATP)悬浮。4℃,12000rpm离心10min,将上清吸出置于新的离心管中。再次离心去掉沉淀。Bradford法测定蛋白浓度,调节至相同浓度。在490μlRDV感病水稻和健康水稻蛋白提取物中分别加入10μl纯化的MBP-IAA10蛋白或10μlMBP蛋白。室温温育。在0min,5min,15min,30min,60min,120min分别取样40μl,加入20μl 3×protein samplebuffer,100℃加热5min,取10μl进行SDS-PAGE。根据预染Marker条带位置,将胶分为三部分,分子量最大的部分(95kDa以上)用来Western blot检测P2蛋白,中间部分(95kDa~72kDa)用来Weestrn blot检测MBP-IAA10,分子量最小的部分(72kDa以下)用考马斯亮蓝染色检测水稻总蛋白的量。RDV-susceptible rice and healthy rice were ground into powder in liquid nitrogen, and an appropriate amount of in vitro degradation buffer (25mM Tris-HCl, pH7.5, 10mM NACl, 10mM MgCl2, 4mM PMSF, 5mM DTT, 10mM ATP) was added to suspend. Centrifuge at 12,000 rpm for 10 min at 4°C, aspirate the supernatant and place it in a new centrifuge tube. Centrifuge again to remove the pellet. The protein concentration was determined by Bradford method and adjusted to the same concentration. 10 μl of purified MBP-IAA10 protein or 10 μl of MBP protein were added to 490 μl of RDV-susceptible rice and healthy rice protein extracts, respectively. Incubation at room temperature. 40μl were sampled at 0min, 5min, 15min, 30min, 60min, and 120min respectively, 20μl of 3×protein sample buffer was added, heated at 100°C for 5min, and 10μl was taken for SDS-PAGE. According to the position of the pre-stained Marker bands, the gel was divided into three parts. The part with the largest molecular weight (above 95kDa) was used to detect P2 protein by Western blot, and the middle part (95kDa-72kDa) was used to detect MBP-IAA10 by Western blot. Parts (below 72kDa) were stained with Coomassie brilliant blue to detect the amount of total rice protein.
结果如图3所示,温育60min后,健康水稻总蛋白提取物中的MBP-IAA10蛋白已经检测不到了,而与RDV感病水稻总蛋白进行温育的MBP-IAA10蛋白直到120min的时候还有一定量的残留。表明MBP-IAA10在RDV感病水稻总蛋白提取物中的稳定性明显高于其在健康水稻总蛋白提取物中的稳定性或者说MBP-IAA10在RDV感病水稻总蛋白提取物中的降解速度明显比健康水稻总蛋白提取物中慢。从MBP组的实验结果可以看出,MBP蛋白在该体系中是稳定存在的,它的量不随时间的变化而变化,说明MBP-IAA10蛋白的降解性质是其中的IAA10蛋白赋予的,因此可以说,MBP-IAA10的降解代表了IAA10的降解。The results are shown in Figure 3. After incubation for 60 min, the MBP-IAA10 protein in the total protein extract of healthy rice was undetectable, while the MBP-IAA10 protein incubated with the total protein of RDV-susceptible rice remained unchanged until 120 min. There is a certain amount of residue. It shows that the stability of MBP-IAA10 in the total protein extract of RDV-susceptible rice is significantly higher than its stability in the total protein extract of healthy rice or the degradation rate of MBP-IAA10 in the total protein extract of RDV-susceptible rice. Significantly slower than in healthy rice total protein extract. It can be seen from the experimental results of the MBP group that the MBP protein exists stably in this system, and its amount does not change with time, indicating that the degradation properties of the MBP-IAA10 protein are endowed by the IAA10 protein, so it can be said that , the degradation of MBP-IAA10 represents the degradation of IAA10.
综上,RDV能够减弱P2的降解。In conclusion, RDV can attenuate the degradation of P2.
三、MBP-IAA10在烟草总蛋白提取物中的降解实验3. Degradation experiment of MBP-IAA10 in tobacco total protein extract
按照2、二、2)中的方法,通过农杆菌注射法分别在烟草叶片中注射含有pCambai1301vector或pCambaia1301-HAS2的农杆菌,表达相应蛋白。将上述叶片分别在液氮中研磨成粉末,加入适量体外降解缓冲液(25mM Tris-HCl,pH7.5,10mM NACl,10mMMgCl2,4mM PMSF,5mM DTT,10mM ATP)悬浮。4℃,12000rpm离心10min,将上清吸出置于新的离心管中。再次离心去掉沉淀。Bradford法测定蛋白浓度,调节至相同浓度。在490μl含有HAP2的和对照的烟草蛋白提取物中分别加入10μl纯化的MBP-IAA10蛋白或10μlMBP蛋白。4℃温育。在0min,20min,40min,80min,180min分别取样40μl,加入20μl 3×proteinsamplebuffer,100℃加热5min,取10μl进行SDS-PAGE。根据预染Marker条带位置,将胶分为三部分,分子量最大的部分(95kDa以上)用来Western blot检测P2蛋白,中间部分(95kDa~72kDa)用来Weestrn blot检测MBP-IAA10,分子量最小的部分(72kDa以下)用考马斯亮蓝染色检测水稻总蛋白的量。According to the method in 2, 2, 2), Agrobacterium containing pCambai1301vector or pCambaia1301-HAS2 was injected into tobacco leaves by Agrobacterium injection method to express the corresponding protein. The above leaves were ground into powders in liquid nitrogen, and an appropriate amount of in vitro degradation buffer (25mM Tris-HCl, pH7.5, 10mM NACl, 10mM MgCl2, 4mM PMSF, 5mM DTT, 10mM ATP) was added to suspend. Centrifuge at 12,000 rpm for 10 min at 4°C, aspirate the supernatant and place it in a new centrifuge tube. Centrifuge again to remove the pellet. The protein concentration was determined by Bradford method and adjusted to the same concentration. 10 μl of purified MBP-IAA10 protein or 10 μl of MBP protein were added to 490 μl of HAP2-containing and control tobacco protein extracts, respectively. Incubation at 4°C. 40μl were sampled at 0min, 20min, 40min, 80min and 180min respectively, 20μl of 3×protein sample buffer was added, heated at 100°C for 5min, and 10μl was taken for SDS-PAGE. According to the position of the pre-stained Marker bands, the gel was divided into three parts. The part with the largest molecular weight (above 95kDa) was used to detect P2 protein by Western blot, and the middle part (95kDa-72kDa) was used to detect MBP-IAA10 by Western blot. Parts (below 72kDa) were stained with Coomassie brilliant blue to detect the amount of total rice protein.
结果如图4所示,温育180min后,对照组烟草总蛋白提取物中的MBP-IAA10蛋白已经检测不到了,而与表达P2的烟草总蛋白进行温育的MBP-IAA10蛋白还有60%的残留。表明MBP-IAA10在含有HAP2的烟草总蛋白提取物中的稳定性明显高于其在不含HAP2的烟草总蛋白提取物中的稳定性或者说MBP-IAA10在P2存在的情况下降解速度明显比没有P2的情况下降解速度慢。从MBP组的实验结果可以看出,MBP蛋白在该体系中是稳定存在的,它的量不随时间的变化而变化,说明MBP-IAA10蛋白的降解性质是其中的IAA10蛋白赋予的,因此可以说,MBP-IAA10的降解代表了IAA10的降解。综上,P2能够阻滞IAA10的降解。The results are shown in Figure 4. After 180 min of incubation, the MBP-IAA10 protein in the tobacco total protein extract of the control group could not be detected, while the MBP-IAA10 protein incubated with the P2-expressing tobacco total protein still had 60%. the residue. It shows that the stability of MBP-IAA10 in tobacco total protein extract containing HAP2 is significantly higher than that in tobacco total protein extract without HAP2, or the degradation rate of MBP-IAA10 in the presence of P2 is significantly higher than that in tobacco total protein extract without HAP2. The degradation rate is slow in the absence of P2. It can be seen from the experimental results of the MBP group that the MBP protein exists stably in this system, and its amount does not change with time, indicating that the degradation properties of the MBP-IAA10 protein are endowed by the IAA10 protein, so it can be said that , the degradation of MBP-IAA10 represents the degradation of IAA10. Taken together, P2 can block the degradation of IAA10.
实施例4、IAA10过量表达能够提高水稻对RDV的敏感性,IAA10RNAi能提高水稻对RDV的抗性Example 4. Overexpression of IAA10 can improve the sensitivity of rice to RDV, and IAA10 RNAi can improve the resistance of rice to RDV
一、表达载体的构建1. Construction of expression vector
1)pCambia1301-FLAGIAA10构建见实施例2、二、1)1) See Example 2, 2, 1 for the construction of pCambia1301-FLAGIAA10
2)pCambia2300-actin-IAA10RNAi构建:以pGAD-IAA10为模版,IAA10-5'SpeI(序列为:5’-GTACTAGTATGAGAGGAGGAGTAGCTGG-3’和IAA10-3'BglII(序列为:5’-GCAGATCTTCAGGATCTGCCTCTTGTTG-3’)进行PCR扩增,得到859bp的PCR产物。将该PCR产物用SpeI和BglII双酶切,连入到用XbalI和BamHI(和SpeI和BglII为同尾酶)酶切过的中间载体pUCCRNAi(未名凯拓公司赠送,公众可从北京大学获得)上,经过测序,重组质粒pUCC-IAA10为将IAA10插入pUCCRNAi质粒的XbalI和BamHI酶切位点间得到的载体。再将用SpeI和BglII双酶切的PCR产物连入到用SpeI和BglII双酶切过pUCC-IAA10质粒中,得到pUCC-IAA10RNAi载体。经过测序,重组质粒pUCC-IAA10RNAi为将IAA10以反向互补形式插入pUCCRNAi质粒的XbalI和BamHI酶切位点和SpeI和BglII酶切位点间得到的载体。2) Construction of pCambia2300-actin-IAA10RNAi: using pGAD-IAA10 as a template, IAA10-5'SpeI (sequence: 5'-GTACTAGTATGAGAGGAGGAGTAGCTGG-3' and IAA10-3'BglII (sequence: 5'-GCAGATCTTCAGGATCTGCCTCTTGTTG-3') Carry out PCR amplification to obtain the PCR product of 859bp.This PCR product is double digested with SpeI and BglII, and is connected to the intermediate vector pUCCRNAi (unnamed) digested with XbalI and BamHI (and SpeI and BglII are homozygous enzymes). Presented by Kaituo Company and available to the public from Peking University), after sequencing, the recombinant plasmid pUCC-IAA10 is a vector obtained by inserting IAA10 between the XbalI and BamHI restriction sites of the pUCCRNAi plasmid. The SpeI and BglII double restriction enzymes The PCR product was connected to the pUCC-IAA10 plasmid that was cut with SpeI and BglII double enzymes to obtain the pUCC-IAA10RNAi vector. After sequencing, the recombinant plasmid pUCC-IAA10RNAi was the XbalI and BamHI enzymes that inserted IAA10 into the pUCCRNAi plasmid with reverse complementation. The resulting vector between the cleavage site and the SpeI and BglII cleavage sites.
将pUCC-IAA10RNAi载体用PstI单酶切,琼脂糖凝胶回收得到1939bp酶切产物,将该产物连接入用PstI单酶切得到的10379bp的pCambia2300-Actin1-ocs载体骨架上,得到重组载体pCambia2300-actin-IAA10RNAi。The pUCC-IAA10RNAi vector was digested with PstI single enzyme, and the 1939bp digested product was recovered from agarose gel, and the product was connected to the 10379bp pCambia2300-Actin1-ocs vector backbone obtained by single enzyme digestion with PstI to obtain the recombinant vector pCambia2300- actin-IAA10RNAi.
二、过量表达转RING1水稻的获得2. Obtaining of overexpressed RING1 rice
1)愈伤组织的诱导培养1) Induction culture of callus
中花11水稻(以下也称为野生型水稻)种子去壳,先用70%乙醇浸泡10min,再用0.1%升汞浸泡30min;进行表面除菌。用大量无菌水洗去种子表面的溶液,用无菌滤纸吸去种子表面的水分。将种子置于成熟胚愈伤诱导培养基平板上,用Parafilm膜封闭平皿边缘,于26℃温箱内避光培养。大约15天后,小心取下长出的愈伤组织,转移到成熟胚继代培养基上,同样条件继续进行培养。每两周需进行一次继代培养。用于转化时,需挑选继代培养5天左右、呈淡黄色的颗粒状愈伤组织。The seeds of
2)农杆菌的培养2) Cultivation of Agrobacterium
将pCambia1301-FLAGIAA10和pCambia2300-actin-IAA10RNAi电转入农杆菌EHA105中,得到重组菌EHA105/pCambia1301-FLAGIAA10和EHA105/pCambia2300-actin-IAA10RNAi。The pCambia1301-FLAGIAA10 and pCambia2300-actin-IAA10RNAi were electroporated into Agrobacterium EHA105 to obtain recombinant strains EHA105/pCambia1301-FLAGIAA10 and EHA105/pCambia2300-actin-IAA10RNAi.
将EHA105/pCambia1301-FLAGIAA10和EHA105/pCambia2300-actin-IAA10RNAi在含有抗生素(50mg/L Kanamycin,50mg/L Rifampicin)的LB平板上划线,28℃培养2天。挑取单菌落接入液体LB培养基中,28℃振荡培养至OD600约为0.5,加入乙酰丁香酮至终浓度100mM,得到用于转化水稻愈伤组织的农杆菌悬液。EHA105/pCambia1301-FLAGIAA10 and EHA105/pCambia2300-actin-IAA10RNAi were streaked on LB plates containing antibiotics (50 mg/L Kanamycin, 50 mg/L Rifampicin) and cultured at 28°C for 2 days. A single colony was picked and placed in liquid LB medium, shaken at 28°C until OD600 was about 0.5, and acetosyringone was added to a final concentration of 100 mM to obtain an Agrobacterium suspension for transforming rice callus.
3)水稻愈伤组织与农杆菌的共培养3) Co-culture of rice callus and Agrobacterium
将继代愈伤组织放入灭过菌的锥形瓶中,倒入农杆菌悬液使之浸没愈伤组织。室温放置20min,并不时轻轻晃动使愈伤组织与菌液充分接触。用无菌的镊子轻轻取出愈伤组织,放于无菌滤纸上吸去多余的菌液,转移到铺有一层无菌滤纸的共培养培养基平板上。28℃暗培养3天,得到经过共培养的愈伤组织。The secondary callus was placed in a sterile Erlenmeyer flask, and the Agrobacterium suspension was poured into it to immerse the callus. Place at room temperature for 20min, and shake gently from time to time to make the callus fully contact with the bacterial liquid. Gently remove the callus with sterile tweezers, place it on sterile filter paper to absorb excess bacterial fluid, and transfer it to a co-cultivation medium plate covered with a layer of sterile filter paper. The cells were cultured in the dark at 28°C for 3 days to obtain the co-cultured callus.
4)抗性愈伤组织的筛选与分化4) Screening and differentiation of resistant callus
将经过共培养的愈伤组织用适量无菌水清洗,除去表面残余的农杆菌,放在筛选培养基上,26℃避光培养进行筛选,两周后转移到新的筛选培养基上继续筛选两周。挑选经过两轮筛选后状态较好的愈伤组织,将其转移到分化培养基平板上,先避光培养3天,然后再转至光照培养箱中(15hr/day)进行光照培养。一个月后可见分化出的小苗。当分化的小苗长至约2cm时,将其转移到锥形瓶中的生根培养基上,继续培养两周左右。选择长势较好、根系发达的小苗,用自来水洗去根部的培养基后移栽入土壤中,收取种子,得到T1代转FLAGIAA10和IAA0RNAi水稻种子,播种得到T1代转FLAGIAA10和IAA0RNAi水稻。Wash the co-cultured callus with an appropriate amount of sterile water to remove the residual Agrobacterium on the surface, put it on the screening medium, culture at 26°C in the dark for screening, and transfer it to a new screening medium two weeks later to continue screening. two weeks. After two rounds of screening, the callus in good condition was selected, transferred to the differentiation medium plate, and cultured in the dark for 3 days, and then transferred to a light incubator (15hr/day) for light culture. The differentiated seedlings can be seen one month later. When the differentiated seedlings grow to about 2 cm, they are transferred to rooting medium in Erlenmeyer flasks and cultured for about two weeks. The seedlings with better growth and developed root system were selected, and the medium from the roots was washed with tap water and then transplanted into the soil, and the seeds were collected to obtain T1 generation transgenic FLAGIAA10 and IAA0RNAi rice seeds, which were sown to obtain T1 generation transgenic FLAGIAA10 and IAA0RNAi rice.
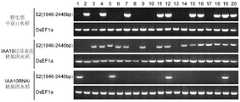
T1代的水稻种子通过潮霉素或G418初步筛选(pCambia1301载体带有潮霉素抗性筛选基因,pCambia2300载体带有G418抗性筛选基因),发芽的种子,表明载体转入水稻。将发芽的种子种入土中,生长2周后,取0.1g叶片,用液氮磨成粉末。The rice seeds of T1 generation were preliminarily screened by hygromycin or G418 (pCambia1301 vector had hygromycin resistance selection gene, pCambia2300 vector had G418 resistance selection gene), and the germinated seeds indicated that the vector was transformed into rice. The germinated seeds were planted in the soil, and after 2 weeks of growth, 0.1 g of leaves were taken and ground into powder with liquid nitrogen.
IAA10过量表达的转基因水稻株系的叶片粉末中加入蛋白提取缓冲液(0.25MTris-Hcl,pH6.8,8%SDS,8%β-巯基乙醇,20%甘油)200μl,冰上孵育10min,100℃煮沸10min,4℃,12000rpm离心10min后取上清,进行SDS-PAGE,转膜之后用Western检测。SDS-PAGE及Western Blot依照公知方法及产品说明书进行。所用抗体为anti-FLAG-HRP(sigma),用抗体anti-Actin检测水稻内院Actin蛋白,作为内参。如图5,在41KDa处出现条带者为阳性,表明基因转入且蛋白表达,选择三个株系#7,#12和#20用于之后的抗病分析实验(图中“-”为野生型中花11水稻,用作负对照;“+”为pCambia1301-FlagIAA10瞬时表达烟草材料所提取蛋白,用作正对照)。Add 200 μl of protein extraction buffer (0.25M Tris-HCl, pH 6.8, 8% SDS, 8% β-mercaptoethanol, 20% glycerol) to the leaf powder of the transgenic rice lines overexpressing IAA10, incubate on ice for 10 min, 100 Boil at ℃ for 10min, centrifuge at 12000rpm for 10min at 4℃, take the supernatant, carry out SDS-PAGE, and use Western detection after transfer to membrane. SDS-PAGE and Western Blot were performed according to known methods and product specifications. The antibody used was anti-FLAG-HRP (sigma), and the antibody anti-Actin was used to detect the Actin protein in the inner courtyard of rice as an internal reference. As shown in Figure 5, the band at 41KDa is positive, indicating gene transfer and protein expression. Three
RNAi的转基因水稻株系的叶片粉末中加入Trizol(Invitrogen),按照说明书所述方法提取RNA。之后,按照下表1,用RQ1Dnase(Promega,货号:M610A)对RNA中的基因组DNA进行消化:Trizol (Invitrogen) was added to the leaf powder of RNAi transgenic rice lines, and RNA was extracted according to the method described in the instructions. Afterwards, the genomic DNA in RNA was digested with RQ1Dnase (Promega, Cat. No. M610A) according to Table 1 below:
表1为消化体系Table 1 shows the digestive system

然后取2ug消化后的RNA进行反转录,具体方法参见invitrogen M-MLV ReverseTranscriptase(货号:28025-021)。将反转得到的cDNA稀释10倍,取4μl进行QuantitativeReal-time PCR反应,引物为qIAA10F:5’-GGTTGCTGGATGGGTGAAGG-3’和qIAA10R:5’-CCTGTCCTCGTAGGTGAGCTGG-3’。反应总体系为20μl,按照SYBR Green Real-Time PCRMaster Mix(TOYOBO)说明书配制。Then take 2ug of digested RNA for reverse transcription. For the specific method, see invitrogen M-MLV ReverseTranscriptase (Cat. No. 28025-021). The cDNA obtained by inversion was diluted 10 times, and 4 μl was taken for QuantitativeReal-time PCR reaction. The primers were qIAA10F: 5'-GGTTGCTGGATGGGTGAAGG-3' and qIAA10R: 5'-CCTGTCCTCGTAGGTGAGCTGG-3'. The total reaction system was 20 μl, prepared according to the instructions of SYBR Green Real-Time PCR Master Mix (TOYOBO).
以OsEF1a作为内参,内参的引物为R10:5’-CTTCCTCTGCGCTTGAGGTG-3’和F12:5‘-ACATTGCCGTCAAGTTTGCTG-3’(参见Zhu et al.,2005.The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases in vivo,leading to reducedbiosynthesis of Gibberellins andrice dwarf symptoms.Plant Physiology.139:1935-1945.)。OsEF1a was used as an internal reference, and the primers of the internal reference were R10: 5'-CTTCCTCTGCGCTTGAGGTG-3' and F12: 5'-ACATTGCCGTCAAGTTTGCTG-3' (see Zhu et al., 2005. The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases in vivo, leading to reduced biosynthesis of Gibberellins andrice dwarf symptoms. Plant Physiology. 139:1935-1945.).
部分株系Real-time PCR鉴定结果如图6所示。大部分IAA10RNAi(图中为节省空间,简写为Ii),转基因株系中IAA10的转录水平下调,为阳性苗。选择IAA10RNAi-1-2(Ii-1-2)、IAA10RNAi-5-1(Ii-5-1)和IAA10RNAi-10(Ii-10-1)三个株系进行后续的侵染实验。The results of Real-time PCR identification of some strains are shown in Figure 6. Most of the IAA10RNAi (space-saving in the figure, abbreviated as Ii), the transcription level of IAA10 in the transgenic lines was down-regulated, and they were positive seedlings. Three lines of IAA10RNAi-1-2(Ii-1-2), IAA10RNAi-5-1(Ii-5-1) and IAA10RNAi-10(Ii-10-1) were selected for subsequent infection experiments.
三、IAA10过量表达能够提高水稻对RDV的敏感性,IAA10RNAi能提高水稻对RDV的抗性3. Overexpression of IAA10 can improve the sensitivity of rice to RDV, and IAA10 RNAi can improve the resistance of rice to RDV
1)通过RT-PCR鉴定RDVS2的表达来鉴定RDV的侵染1) Identify the infection of RDV by identifying the expression of RDVS2 by RT-PCR
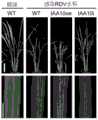
用携带RDV(参见Zhu et al.,2005.The Rice Dwarf Virus P2Proteininteractswith ent-Kaurene Oxidases in vivo,leading to reduced biosynthesis ofGibberellins andrice dwarf symptoms.Plant Physiology.139:1935-1945.公众可从北京大学获得。)的叶蝉(病原菌为水稻矮缩病毒(Rice Dwarf Virus))接种T1代转IAA10过量表达和RNAi转基因水稻和野生型水稻中花11,每种水稻接种20株,白天30度,晚上22度,湿度60%培养,每株接5头叶蝉,咬食三天后将叶蝉捉出,咬食过的水稻在阳光温室中培养(自然光,温度。实验重复3次,结果取平均值)。with RDV (see Zhu et al., 2005. The Rice Dwarf Virus P2Protein interacts with ent-Kaurene Oxidases in vivo, leading to reduced biosynthesis of Gibberellins andrice dwarf symptoms. Plant Physiology. 139:1935-1945. Publicly available from Peking University.) The leafhoppers (the pathogen is Rice Dwarf Virus) were inoculated with T1 generation transgenic IAA10 overexpression and RNAi transgenic rice and wild-
接毒4周后,提取20株T1代转IAA10过量表达、20株RNAi转基因水稻和20株野生型水稻中花11总RNA,之后,按照表1,用RQ1Dnase(Promega,货号:M610A)对RNA中的基因组DNA进行消化,然后取2ug消化后的RNA进行反转录和半定量PCR,具体方法参见invitrogen M-MLV Reverse Transcriptase(货号:28025-021),扩增产物为RDV P2部分蛋白的编码序列S2;其中使用的引物为:RDVS2F1946:CTATACCACGATGAAGGCCT和RDVS2R2446:TAATGGTACCGATGGTTACC。得到500bp大小条带的为感染RDV阳性,没有的为感染RDV阴性。After 4 weeks of inoculation, the total RNA of
以OsEF1a作为内参,内参的引物为R10:5’-CTTCCTCTGCGCTTGAGGTG-3’和F12:5‘-ACATTGCCGTCAAGTTTGCTG-3’(参见Zhu et al.,2005.The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases in vivo,leading to reducedbiosynthesis of Gibberellins andrice dwarf symptoms.Plant Physiology.139:1935-1945.)。结果如图7所示,可以看出,20株IAA10过量表达T1代水稻中有15株扩增得到500bp的S2;20株IAA10RNAi转基因水稻中有4株扩增到500bp的S2;20株野生型水稻中花11中有10株扩增得到500bp的S2;IAA10过量表达能够提高水稻对RDV的敏感性,IAA10RNAi能提高水稻对RDV的抗性。OsEF1a was used as an internal reference, and the primers of the internal reference were R10: 5'-CTTCCTCTGCGCTTGAGGTG-3' and F12: 5'-ACATTGCCGTCAAGTTTGCTG-3' (see Zhu et al., 2005. The Rice Dwarf VirusP2Proteininteracts with ent-Kaurene Oxidases in vivo, leading to reduced biosynthesis of Gibberellins andrice dwarf symptoms. Plant Physiology. 139:1935-1945.). The results are shown in Figure 7. It can be seen that 15 of the 20 IAA10-overexpressing T1 rice plants were amplified to obtain 500 bp S2; 4 of the 20 IAA10 RNAi transgenic rice plants were amplified to 500 bp S2; 20 were wild-
2)通过表型确定RDV侵染率2) Determination of RDV infection rate by phenotype
接毒4周后,观察20株T1代转IAA10过量表达、20株RNAi转基因水稻和20株野生型水稻中花11的症状(其中感染RDV病毒的为植株矮缩,叶色浓绿,叶片僵硬,在叶片或叶鞘上出现白色斑点并与叶脉平行排列成虚线状,无感染RDV病毒的为叶片或叶鞘上不出现白色斑点),统计有症状植株数目,计算带毒率=(有表型株数/总株数)*%。After 4 weeks of inoculation, observe the symptoms of 20 T1 generation transgenic IAA10 overexpression, 20 RNAi transgenic rice and 20 wild-type rice Zhonghua 11 (wherein those infected with RDV virus are dwarf plants, dark green leaves, and Stiff, white spots appear on the leaves or leaf sheaths and are arranged in dashed lines parallel to the leaf veins, no white spots appear on the leaves or leaf sheaths without RDV virus infection), count the number of symptomatic plants, and calculate the virus-carrying rate = (with phenotype Number of plants/total number of plants)*%.
结果统计如下表2:The statistics of the results are as follows in Table 2:
表2为转基因水稻在病毒侵染后带毒率统计结果Table 2 shows the statistical results of the virus-carrying rate of transgenic rice after virus infection

可以看出,IAA10过量表达水稻感病率更高,而IAA10RNAi水稻感病率相对较低。除此以外,我们对不同株系感病水稻进行了拍照,如图8所示,为侵染后4周健康,IAA10过量表达和IAA10RNAi转基因水稻的感病症状图。可以看出,IAA10过量表达的水稻感病症状更强,表现为矮化程度更强,叶片上表示病毒积累的斑点数量更多;而IAA10RNAi的转基因水稻的感病症状则相对更弱,表现为矮化程度相对野生型更弱,而病斑数量更少(图8,图中WT代表野生型中花11水稻,IAA10oe代表IAA10过量表达转基因水稻,IAA10i代表IAA10RNAi的转基因水稻)。与野生型水稻相比,IAA10过量表达转基因水稻更加易感,而IAA10RNAi转基因水稻更加抗病。It can be seen that the IAA10 overexpressed rice has a higher susceptibility rate, while the IAA10RNAi rice susceptibility rate is relatively low. In addition, we took pictures of the susceptible rice lines of different lines, as shown in Figure 8, which is a graph of the disease symptoms of healthy, IAA10 overexpressed and IAA10RNAi




Claims (1)
Translated fromChinesePriority Applications (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201510882608.6ACN106831967B (en) | 2015-12-03 | 2015-12-03 | Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus |
Applications Claiming Priority (1)
| Application Number | Priority Date | Filing Date | Title |
|---|---|---|---|
| CN201510882608.6ACN106831967B (en) | 2015-12-03 | 2015-12-03 | Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus |
Publications (2)
| Publication Number | Publication Date |
|---|---|
| CN106831967A CN106831967A (en) | 2017-06-13 |
| CN106831967Btrue CN106831967B (en) | 2020-08-04 |
Family
ID=59149731
Family Applications (1)
| Application Number | Title | Priority Date | Filing Date |
|---|---|---|---|
| CN201510882608.6AActiveCN106831967B (en) | 2015-12-03 | 2015-12-03 | Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus |
Country Status (1)
| Country | Link |
|---|---|
| CN (1) | CN106831967B (en) |
Families Citing this family (3)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN112390866B (en)* | 2019-08-14 | 2022-07-15 | 北京大学 | Application of OsARF12 gene in improving resistance of rice to rice dwarf virus |
| CN111253480B (en)* | 2020-03-04 | 2021-08-06 | 宁波大学 | Rice transcription factor OsARF17 gene and its application in plant breeding for resistance to black-streaked dwarf virus |
| CN114958906B (en)* | 2022-06-22 | 2023-08-08 | 贵州省烟草公司毕节市公司 | Genes, promoters and applications related to low potassium stress in tobacco |
Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN102224247A (en)* | 2008-09-24 | 2011-10-19 | 巴斯夫植物科学有限公司 | Plants having enhanced yield-related traits and a method for making the same |
| CN103194431A (en)* | 2013-04-17 | 2013-07-10 | 北京大学 | Application of RING1 protein for improving RDV (rice dwarf virus) resistance of plants |
- 2015
- 2015-12-03CNCN201510882608.6Apatent/CN106831967B/enactiveActive
Patent Citations (2)
| Publication number | Priority date | Publication date | Assignee | Title |
|---|---|---|---|---|
| CN102224247A (en)* | 2008-09-24 | 2011-10-19 | 巴斯夫植物科学有限公司 | Plants having enhanced yield-related traits and a method for making the same |
| CN103194431A (en)* | 2013-04-17 | 2013-07-10 | 北京大学 | Application of RING1 protein for improving RDV (rice dwarf virus) resistance of plants |
Non-Patent Citations (2)
| Title |
|---|
| Transcriptome profiling confirmed correlations between symptoms and transcriptional changes in RDV infected rice and revealed nucleolus as a possible target of RDV manipulation;Liang Yang 等;《Virology Journal》;20140506;第11卷;第1-10页* |
| 水稻矮缩病毒对3种内源激素含量及代谢相关基因转录水平的影响;吴建国 等;《植物病理学报》;20100228;第40卷(第2期);第151-158页* |
Also Published As
| Publication number | Publication date |
|---|---|
| CN106831967A (en) | 2017-06-13 |
Similar Documents
| Publication | Publication Date | Title |
|---|---|---|
| CN101942449A (en) | Methods of increasing abiotic stress tolerance and/or biomass in plants and plants generated thereby | |
| CN106244599B (en) | A kind of Panax notoginseng disease course related protein 1 family gene PnPR1-2 and its application | |
| CN103194456B (en) | Lilium regale antifungal gene Lr14-3-3 and application thereof | |
| CN103194431B (en) | Application of RING1 protein for improving RDV (rice dwarf virus) resistance of plants | |
| WO2016065896A1 (en) | Applications of osago18 protein or coding gene thereof in regulating resistance of plant against rice dwarf virus or viruses of same family | |
| CN105755020A (en) | Radix notoginseng mitogen-activated protein kinase kinase gene PnMAPKK1 and application thereof | |
| CN106831967B (en) | Application of reducing IAA10 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus | |
| US10041086B2 (en) | Method for production of transgenic cotton plants | |
| CN113980986B (en) | Application of CRK22 gene and encoding protein thereof in potato stress-resistant breeding | |
| CN104878019A (en) | Yangbi walnut germin-like protein gene JsGLP1 and application thereof | |
| Zaulda et al. | A cowpea severe mosaic virus-based vector simplifies virus-induced gene silencing and foreign protein expression in soybean | |
| CN107267526A (en) | Pseudo-ginseng myb transcription factor gene PnMYB2 and its application | |
| CN103937819B (en) | A kind of lilium regale wilson glutathione S-transferase gene LrGSTL1 and application thereof | |
| CN102899333B (en) | Rice salt stress related gene SIDP364 and coding protein and application thereof | |
| CN117625684B (en) | Salvia SmWRKY protein and application of coding gene thereof in regulation and control of salt tolerance of plants | |
| Tiwari et al. | A tau class GST, OsGSTU5, interacts with VirE2 and modulates the Agrobacterium-mediated transformation in rice | |
| CN116970621B (en) | Salvia miltiorrhizae stress-resistance related protein SmDREB2A and encoding gene and application thereof | |
| CN103320448B (en) | Lilium regle bZIP transcription factor LrbZIP1 and application | |
| CN114591409B (en) | Application of TaDTG6 Protein in Improving Plant Drought Resistance | |
| KR102674984B1 (en) | CaSIRF1 gene and Method for improving the resistance to the drought stress using CaSIRF1 in plants | |
| CN101955519B (en) | Plant DREB transcription factor, coding gene thereof and use thereof | |
| CN104945492A (en) | Plant stress tolerance associated protein TaAREB3 as well as encoding gene and application thereof | |
| CN110042113B (en) | Rice grain shape positive regulator gene OsMAPKKK70, its encoded protein and its application | |
| CN112390866B (en) | Application of OsARF12 gene in improving resistance of rice to rice dwarf virus | |
| CN109295024B (en) | Application of OsSAMS1 protein and coding gene expression thereof in improving resistance of plants to rice dwarf virus |
Legal Events
| Date | Code | Title | Description |
|---|---|---|---|
| PB01 | Publication | ||
| PB01 | Publication | ||
| SE01 | Entry into force of request for substantive examination | ||
| SE01 | Entry into force of request for substantive examination | ||
| GR01 | Patent grant | ||
| GR01 | Patent grant |