Introduction
The endocannabinoid system (ECS) is composed of G protein-coupled cannabinoid receptors, namely cannabinoid receptor-1 (CB1) and cannabinoid receptor-2 (CB2)1,2; the endogenous cannabinoids called endocannabinoids, such as the lipids anandamide and 2-arachidonoylglycerol3,4; and the enzymes involved in their synthesis and inactivation5. The family of endocannabinoids has recently grown to include a group of peptide ligands (so-called pepcans) and other lipid molecules, such as lipoxin and pregnenolone, interestingly acting as allosteric enhancers or signal-specific inhibitors (SSIs) of CB1 receptors6.
One of the main characteristics of the ECS is its broad distribution throughout the body. In this review, we will specifically focus our attention on the CB1 receptor-dependent functions in the nervous system (particularly the brain). The CB1 receptor is considered the most abundant metabotropic receptor in the brain7. It was cloned in 19901 and its distribution has been well characterized in both rodents8,9 and humans10. These receptors are particularly rich in the central nervous system11,12, where they control a wide spectrum of physiological and pathological conditions, including brain development, learning and memory, motor behavior, regulation of appetite, body temperature, pain perception, inflammation, and they are involved in various psychiatric, neurological, and neurodevelopmental disorders13–17.
This review highlights recent findings that challenge or extend accepted “dogmas” of CB1 receptor signaling. Thus, it discusses where CB1 receptors are localized, the importance of CB1 receptors outside the brain, and new strategies to pharmacologically act on these receptors. Importantly, the understanding of where, which, and how CB1 receptor function is mandatory to improve the pharmacological strategies to act on this promising therapeutic target.
Localization of CB1 receptors in different neuronal types
CB1 receptor localization has been widely studied during the last few decades18. Thus, early studies provided strong evidence for a presynaptic localization of CB1 receptors, from where they can control the neurotransmitter release7,19. However, the somatodendritic localization of CB1 receptors cannot be discarded, as processes of self-inhibition through these receptors have been demonstrated in the cortex20–23. According to this, recent work describes that somatodendritic CB1 receptors control a specific postsynaptic signaling cascade important for the cognitive impairment induced by cannabinoids24. Therefore, more studies are needed to clarify the relative involvement of pre- or post-synaptic CB1 receptors in brain functions and how this can affect our general view of how the ECS controls synaptic transmission.
Interestingly, new experimental approaches (e.g., imaging tools) have shown the expression of CB1 receptors in different neuronal types, including GABAergic, glutamatergic, and serotonergic neurons, among others8,25–28. Moreover, although the anatomical presence of CB1 receptors in cholinergic, noradrenergic, or dopaminergic neurons has not been fully characterized, cannabinoids are known to control acetylcholine and dopamine release29,30. For example, it has been recently shown that CB1 receptors can specifically control cholinergic over glutamatergic transmission at single synapses that co-release both neurotransmitters31.
Importantly, the expression levels of CB1 receptors can drastically differ among different cell types and can diverge between different brain regions12,25,32,33. This widely distributed and differential expression in the brain reflects the complexity, and can explain the variety of functions, of the ECS. For instance, this specific distribution can explain some of the bimodal effects of cannabinoid drugs34,35. Thus, recent studies demonstrated how CB1 receptors localized in GABAergic neurons can control food intake34, running related behaviors36,37, drug addiction38,39, and learning and memory processes40,41, among other behaviors, whereas CB1 receptors localized in glutamatergic neurons control neuroprotection42, olfactory processes25, fear memories43, social behaviors44, and anxiety35, among others. Moreover, CB1 receptors present in serotonergic neurons can modulate emotional responses45.
Localization of CB1 receptors in other cell types or intracellular organelles
The biased neuron-centric view in the ECS field changed when CB1 receptors were found in another type of brain cells, the glial cells46–49. Moreover, recent studies have demonstrated how the astroglial CB1 receptor can modulate important physiological functions in behavior and synaptic plasticity such as learning and memory and long-term depression in the hippocampus50–52. Therefore, this receptor can shape synaptic transmission via astroglial signaling53. By doing this, it modulates the effects of exogenous cannabinoids on working memory46 and, notably, can also determine the selective activity of specific circuits in the striatum54. Thus, the improvement of the current tools will consolidate this knowledge to better elucidate the role of CB1 receptors and astrocytes on brain functioning55. Interestingly, recent findings have shown how CB1 receptors can modulate microglia activation, suggesting its presence in this cell type49.
Although CB1 receptors are localized primarily at the plasma membrane, more and more evidence suggests the presence of functional intracellular CB1 receptors56,57. For instance, a portion of these receptors is functionally present in cell mitochondria58. In the past, previous data showed that cannabinoids can alter mitochondrial functions, but these effects were fully ascribed to unspecific membrane disturbance induced by these lipid molecules59,60. However, recent results challenge this idea, indicating that CB1 receptors are also present in mitochondrial membranes in the periphery, such as in spermatozoa61 or skeletal muscles62, and in the brain, where they directly regulate mitochondrial oxidative phosphorylation (OXPHOS) activity58,63,64 or can impact feeding behavior65. However, further studies and more direct, specific, and powerful tools are needed to investigate the role of mitochondrial or other intracellular CB1 receptors on synaptic transmission, brain functions, and behavior. Interestingly, brain mitochondrial functions have been recently causally associated to anxiety-related responses in the nucleus accumbens66, demonstrating how brain energetics can impact behavior.
Localization of CB1 receptors in the periphery
In the last two decades, CB1 receptors have been described in a number of peripheral tissues, including fat tissue67, gastrointestinal tract68, mouth and oral cavity69, eye70, cardiovascular system71, liver72, pancreas73, immune system74, bone75, skin76, and skeletal muscle77. Indeed, it seems that the ECS is present in a large majority of tissues and its specific functions have recently been investigated78.
The complex interactions between peripheral organs and the central nervous system raised a particular interest within the neuroscience field. In this sense, it is worth discussing how the peripheral processes modulated by the CB1 receptors are affecting the central nervous system functions. A recent study demonstrated that the peripheral sympathetic activity controlled by CB1 receptors is necessary for central functions, such as hypophagia and anxiety-like effects79. Other potential examples of the roles of CB1 receptors in the periphery-brain connection are the control of the release of stress hormones from the adrenal glands80 or the modulation of gut functions impacting on behavioral responses. Indeed, a close interaction between adipose tissue, gut bacteria, and the endocannabinoid system has been proposed in the context of obesity81,82.
New advances in the CB1 receptor pharmacology
Several orthosteric ligands of CB1 receptors have been described in the last few decades, including natural or synthetic CB1 receptor agonists (e.g., Δ9-tetrahydrocannabinol [THC], CP-55,940), antagonists (e.g., rimonabant), and orthosteric endocannabinoids6,83. Moreover, endocannabinoids seem also to target non-cannabinoid receptors (e.g., G protein-coupled receptor 55 receptors)84,85 and ion channels (e.g., serotonergic, nicotinic acetylcholine receptors, or vanilloid receptors)86, particularly at concentrations at which they have been found to interact with CB1 or CB2 receptors6,87. Notably, the orthosteric action of CB1 receptor agonists and antagonists induces important side effects88,89. For example, rimonabant, known as a partial antagonist/inverse agonist, showed different side effects in humans88. In this sense, different strategies have been shown to improve the safety profile and overcome the side effects induced by CB1 antagonists, such as the neutral CB1 antagonists90.
Interestingly, the pharmacology of CB1 receptors is nowadays also focused in the recent developments on putative allosteric binding sites of these receptors and how this can be translated into new therapeutic approaches. As cannabinoid ligands present an interesting therapeutic profile91, the development of new and safer drugs such as CB1 receptor allosteric modulators is needed. Indeed, this strategy has become a hot topic in the G protein-coupled receptors field and there are different positive and negative allosteric modulators described (PAMs and NAMs, respectively)92,93. Consequently, different compounds have been developed as exogenous CB1 allosteric modulators, including the indole derivatives (e.g., the NAM “ORG” compounds)94, urea derivatives (e.g., the NAM PSNCBAM-1)95, and other small molecules that also display a PAM profile, such as RTI-37196. Importantly, recent work also identified natural PAMs and NAMs of CB1 receptors, such as the lipoxin A4, the hemopressin pepcan-12, and pregnenolone97,98, which might represent model chemical structures for the development of new drugs. Although numerous studies have fully characterized the chemical and signaling properties of these new synthetic or natural compounds97,98, thein vivo effects of all these drugs modulating physiological or pathological conditions constitutes an emerging area in the cannabinoid field. In this context, the neurosteroid pregnenolone exerts peculiar effects on CB1 receptor signaling. Indeed, pregnenolone, by binding to a specific identified site on CB1 receptors, displays an interesting SSI profile: whereas CB1-dependent modulation of cytoplasmic cyclic AMP signaling is unaltered by pregnenolone, the neurosteroid fully blocks the activation of extracellularly regulated kinases (ERKs) and the inhibition of mitochondrial activity by cannabinoids63. By these mechanisms, the SSI pregnenolone blocks different central effects of THC, including memory impairment, hypolocomotion, and cannabinoid self-administration in rodents63. Other compounds have been shown to alter CB1 receptor-dependent effects. For instance, the synthetic PAM ZCZ011 reduces neuropathic pain99, whereas the PAM lipoxin A4 shows anti-inflammatory effects100. Interestingly, it was recently shown that cannabidiol, which has been previously reported as a CB1 receptor antagonist, behaves also as a non-competitive NAM of CB1 receptors, despite its low affinity to these receptors101.
The allosteric modulators of CB1 receptors are not the only therapeutic agents recently proposed. Indeed, the effects of several phytocannabinoids in preclinical models of central nervous system diseases and, where available, clinical trials have been investigated, suggesting a promising phytocannabinoid-based medicine102. Another factor that can change the CB1 receptor pharmacology is heteromerization with other receptors. Heteromers of CB1 receptors and other proteins recently emerged as an important target of thein vivo effects of cannabinoids103–105. Notably, these heterocomplexes could be potentially modulated104 and this implies another pharmacological tool to act on CB1 receptor signaling. Moreover, present evidence points to the membrane environment as another critical regulator of CB1 receptor signaling, and this can be potentially exploited for the development of novel therapeutic compounds106. Finally, a G protein-coupled receptor such as the CB1 receptor may also have a constitutive, ligand-free mode of signaling, as has been shown in hippocampal GABAergic synapses107. All of these new ideas demonstrate that the research community may dedicate more effort to tackle CB1 receptors.
Conclusions
This short review focused on the new findings in CB1 receptor research. However, the ECS comprises other components such as CB2 receptors, the endocannabinoids, and the enzymes responsible for their synthesis and degradation. In this sense, recent advances have demonstrated the importance of CB2 receptors in the brain108–110, the presence of other endocannabinoid-like molecules111,112, other potential receptors that can be activated by endocannabinoids87, and interesting findings regarding the localization and pharmacology of the enzymes involved in the metabolism of these endocannabinoids113,114. In brief, the actual picture of how the endocannabinoid system works is quite complicated and more efforts are needed to try to merge the old and the new ideas in this field (Figure 1).
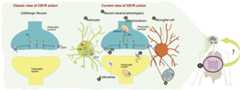
Figure 1. Schematic comparison between the classic and the current view of the CB1 receptor functional expression.
On the left panel, the classic view of the CB1 receptor is represented. The CB1 receptor was thought to be exclusively localized in GABAergic neurons, where it was demonstrated to inhibit neurotransmitter release. On the right panel, the current view of the CB1 receptor is illustrated. Different advances have completely changed this picture: (A) The CB1 receptor is present in different neuronal types and in glial cells, both in astrocytes (B) and potentially in microglia (C). Furthermore, it is found intracellularly in the mitochondria (D) and endosomes (E). The view of a canonical retrograde system changed after the CB1 receptor localization in postsynaptic somatodentritic neurons was demonstrated (F). Nowadays, we know that CB1 receptor presents allosteric binding sites (G) and that it could form heteromers (H). Beyond the brain, the CB1 receptor is widely expressed in the periphery (I), where it can modulate the periphery-brain connection. All of this new knowledge reflects the complexity of the central nervous system and the advance in neuroscience, positing the CB1 receptor as an ideal tool for studying brain functions. CB1, cannabinoid receptor-1; CB2, cannabinoid receptor-2; eCB, endocannabinoid; NT, neurotransmitter.
An open question in the cannabinoid field is whether the cellular diversity of CB1 functions could improve the therapeutic exploitation of cannabinoid-based drugs. One can speculate whether different CB1ligands can mediate different signaling pathways by selectively controlling different CB1 receptors present in different cellular populations. Likewise, it is possible that specific drugs could target exclusively mitochondrial CB1 (mtCB1) receptors or could avoid activation of intracellular pools of CB1. More studies will be needed to answer these questions, but there is already some evidence demonstrating a different pharmacological profile between CB1 receptors expressed in GABAergic and glutamatergic cells. Thus, “glutamatergic” CB1 receptors are more sensitive to low doses of agonists and are endowed with stronger intracellular coupling, whereas “GABAergic” pools of the receptor are activated by higher doses of agonists and produce lower activation of G proteins34,35,43,115. Therefore, one could speculate that specific compounds able to selectively activate different cellular subpopulations of CB1 receptors could be developed. Moreover, combinations of drugs able to modulate glutamatergic or GABAergic neurotransmission with cannabinoid agonists have been shown to promote specific effects of CB1 receptors and inhibit others116. It is also interesting to note that both perisomatic and dendritic GABAergic synapses use phasic endocannabinoid signaling, but the tonic form of cannabinoid signaling is present only in perisomatic cells107. Moreover, a recent study80 shows that the peptide endocannabinoids, known as pepcans, act as endogenous allosteric modulators of CB1 activity exclusively on noradrenergic neurons, demonstrating a cell type-specific regulatory role on endocannabinoid signaling. All of these new and exciting findings suggest that the better we understand cannabinoid signaling, the closer we are to developing specific and local pharmacological drugs that may have importance in brain disorders.
Overall, the new and exciting findings suggesting different and specific localizations of the ECS components and the new strategies proposed to tackle their activity of this receptor open the door to new questions (Table 1). Indeed, the endocannabinoid system has been related to many physiological and pathological functions13,18,117, and the better understanding of these new evidences will bring more light to exploit the therapeutically beneficial properties of this widely spread neuromodulator system in the brain and in the body.
Table 1. Open questions in the cannabinoid receptor-1 (CB1) receptor field.
| Open questions in the endocannabinoid field. |
|---|
Is the cell type-specific CB1 receptor signaling an open door to
develop new therapeutic tools? |
Is the endocannabinoid system exclusively a retrograde
neuromodulator system? |
How is the subcellular CB1 receptor distributed in the different
cell types? |
| How can CB1 receptors control neurotransmitter co-release? |
Which physiological and pathological functions are modulated
by intracellular CB1 receptors? |
Is there specific or differential CB2 receptor expression in
different cell types? |
Is the allosteric modulation of CB1 receptors a good therapeutic
approach for pathological conditions? |
Will it be possible to create compounds that target CB1 receptors
in specific cell types or subcellular localizations? |
Abbreviations
CB1, cannabinoid receptor-1; CB2, cannabinoid receptor-2; ECS, endocannabinoid system; NAM, negative allosteric modulator; PAM, positive allosteric modulator; SSI, signal-specific inhibitor; THC, Δ9-tetrahydrocannabinol.
Competing interests
The authors declare that they have no competing interests.
Grant information
We thank all the members of the GM lab for useful discussions. This work was supported by INSERM (to GM), EU-FP7 (PAINCAGE, HEALTH-603191 to GM and FP7-PEOPLE-2013-IEF-623638 to AB-G), European Research Council (Endofood, ERC-2010-StG-260515; CannaPreg, ERC-2014-PoC-640923, to GM), Fondation pour la Recherche Medicale (DRM20101220445, to GM and LB), Human Frontiers Science Program (to GM), Region Aquitaine (to GM), French State/Agence Nationale de la Recherche/LabEx BRAIN (ANR-10-LABX-0043 to GM), Fyssen Foundation (to ES-G), CONACyT (to ES-G), French State/Agence Nationale de la Recherche/IdEx (ANR-10-IDEX-03-02 to AB-G), and French State/Agence Nationale de la Recherche/Blanc (NeuroNutriSens ANR-13-BSV4-0006-02 to GM).
Faculty Opinions recommendedReferences
- 1. Matsuda LA, Lolait SJ, Brownstein MJ, et al.: Structure of a cannabinoid receptor and functional expression of the cloned cDNA.Nature. 1990;346(6284): 561–4.PubMed Abstract |Publisher Full Text
- 2. Howlett AC, Barth F, Bonner TI, et al.: International Union of Pharmacology. XXVII. Classification of cannabinoid receptors.Pharmacol Rev. 2002;54(2): 161–202.PubMed Abstract
- 3. Little PJ, Compton DR, Johnson MR, et al.: Pharmacology and stereoselectivity of structurally novel cannabinoids in mice.J Pharmacol Exp Ther. 1988;247(3): 1046–51.PubMed Abstract
- 4. Devane WA, Hanus L, Breuer A, et al.: Isolation and structure of a brain constituent that binds to the cannabinoid receptor.Science. 1992;258(5090): 1946–9.PubMed Abstract |Publisher Full Text
- 5. Pacher P, Bátkai S, Kunos G: The endocannabinoid system as an emerging target of pharmacotherapy.Pharmacol Rev. 2006;58(3): 389–462.PubMed Abstract |Publisher Full Text |Free Full Text
- 6. Pertwee RG: Endocannabinoids and Their Pharmacological Actions.Handb Exp Pharmacol. 2015;231: 1–37.PubMed Abstract |Publisher Full Text
- 7. Kano M, Ohno-Shosaku T, Hashimotodani Y, et al.: Endocannabinoid-mediated control of synaptic transmission.Physiol Rev. 2009;89(1): 309–80.PubMed Abstract |Publisher Full Text
- 8. Herkenham M, Lynn AB, Johnson MR, et al.: Characterization and localization of cannabinoid receptors in rat brain: a quantitativein vitro autoradiographic study.J Neurosci. 1991;11(2): 563–83.PubMed Abstract
- 9. Tsou K, Brown S, Sañudo-Peña MC, et al.: Immunohistochemical distribution of cannabinoid CB1 receptors in the rat central nervous system.Neuroscience. 1998;83(2): 393–411.PubMed Abstract |Publisher Full Text
- 10. Westlake TM, Howlett AC, Bonner TI, et al.: Cannabinoid receptor binding and messenger RNA expression in human brain: anin vitro receptor autoradiography andin situ hybridization histochemistry study of normal aged and Alzheimer's brains.Neuroscience. 1994;63(3): 637–52.PubMed Abstract |Publisher Full Text
- 11. Hu SS, Mackie K: Distribution of the Endocannabinoid System in the Central Nervous System.Handb Exp Pharmacol. 2015;231: 59–93.PubMed Abstract |Publisher Full Text
- 12. Freund TF, Katona I, Piomelli D: Role of endogenous cannabinoids in synaptic signaling.Physiol Rev. 2003;83(3): 1017–66.PubMed Abstract |Publisher Full Text
- 13. Katona I, Freund TF: Multiple functions of endocannabinoid signaling in the brain.Annu Rev Neurosci. 2012;35: 529–58.PubMed Abstract |Publisher Full Text |Free Full Text
- 14. Heifets BD, Castillo PE: Endocannabinoid signaling and long-term synaptic plasticity.Annu Rev Physiol. 2009;71: 283–306.PubMed Abstract |Publisher Full Text |Free Full Text
- 15. Mechoulam R, Parker LA: The endocannabinoid system and the brain.Annu Rev Psychol. 2013;64: 21–47.PubMed Abstract |Publisher Full Text
- 16. Lu H, Mackie K: An Introduction to the Endogenous Cannabinoid System.Biol Psychiatry. 2016;79(7): 516–25.PubMed Abstract |Publisher Full Text |Free Full Text
- 17. Lutz B, Marsicano G, Maldonado R, et al.: The endocannabinoid system in guarding against fear, anxiety and stress.Nat Rev Neurosci. 2015;16(12): 705–18.PubMed Abstract |Publisher Full Text
- 18. Busquets-Garcia A, Desprez T, Metna-Laurent M, et al.: Dissecting the cannabinergic control of behavior: Thewhere matters.Bioessays. 2015;37(11): 1215–25.PubMed Abstract |Publisher Full Text
- 19. Ohno-Shosaku T, Tanimura A, Hashimotodani Y, et al.: Endocannabinoids and retrograde modulation of synaptic transmission.Neuroscientist. 2012;18(2): 119–32.PubMed Abstract |Publisher Full Text
- 20. Bacci A, Huguenard JR, Prince DA: Long-lasting self-inhibition of neocortical interneurons mediated by endocannabinoids.Nature. 2004;431(7006): 312–6.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 21. Min R, Testa-Silva G, Heistek TS, et al.: Diacylglycerol lipase is not involved in depolarization-induced suppression of inhibition at unitary inhibitory connections in mouse hippocampus.J Neurosci. 2010;30(7): 2710–5.PubMed Abstract |Publisher Full Text
- 22. Marinelli S, Pacioni S, Bisogno T, et al.: The endocannabinoid 2-arachidonoylglycerol is responsible for the slow self-inhibition in neocortical interneurons.J Neurosci. 2008;28(50): 13532–41.PubMed Abstract |Publisher Full Text |Free Full Text
- 23. Marinelli S, Pacioni S, Cannich A, et al.: Self-modulation of neocortical pyramidal neurons by endocannabinoids.Nat Neurosci. 2009;12(12): 1488–90.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 24. Maroso M, Szabo GG, Kim HK, et al.: Cannabinoid Control of Learning and Memory through HCN Channels.Neuron. 2016;89(5): 1059–73.PubMed Abstract |Publisher Full Text |Free Full Text
- 25. Soria-Gómez E, Bellocchio L, Reguero L, et al.: The endocannabinoid system controls food intake via olfactory processes.Nat Neurosci. 2014;17(3): 407–15.PubMed Abstract |Publisher Full Text
- 26. Marsicano G, Lutz B: Neuromodulatory functions of the endocannabinoid system.J Endocrinol Invest. 2006;29(3 Suppl): 27–46.PubMed Abstract
- 27. Kawamura Y, Fukaya M, Maejima T, et al.: The CB1 cannabinoid receptor is the major cannabinoid receptor at excitatory presynaptic sites in the hippocampus and cerebellum.J Neurosci. 2006;26(11): 2991–3001.PubMed Abstract |Publisher Full Text
- 28. Häring M, Enk V, Aparisi Rey A, et al.: Cannabinoid type-1 receptor signaling in central serotonergic neurons regulates anxiety-like behavior and sociability.Front Behav Neurosci. 2015;9: 235.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 29. Hernandez G, Cheer JF: To Act or Not to Act: Endocannabinoid/Dopamine Interactions in Decision-Making.Front Behav Neurosci. 2015;9: 336.PubMed Abstract |Publisher Full Text |Free Full Text
- 30. Martin HG, Bernabeu A, Lassalle O, et al.: Endocannabinoids Mediate Muscarinic Acetylcholine Receptor-Dependent Long-Term Depression in the Adult Medial Prefrontal Cortex.Front Cell Neurosci. 2015;9: 457.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 31. Soria-Gómez E, Busquets-Garcia A, Hu F, et al.: Habenular CB1 Receptors Control the Expression of Aversive Memories.Neuron. 2015;88(2): 306–13.PubMed Abstract |Publisher Full Text
- 32. Mackie K: Distribution of cannabinoid receptors in the central and peripheral nervous system.Handb Exp Pharmacol. 2005; (168): 299–325.PubMed Abstract |Publisher Full Text
- 33. Katona I: Cannabis and Endocannabinoid Signaling in Epilepsy.Handb Exp Pharmacol. 2015;231: 285–316.PubMed Abstract |Publisher Full Text
- 34. Bellocchio L, Lafenêtre P, Cannich A, et al.: Bimodal control of stimulated food intake by the endocannabinoid system.Nat Neurosci. 2010;13(3): 281–3.PubMed Abstract |Publisher Full Text
- 35. Rey AA, Purrio M, Viveros MP, et al.: Biphasic effects of cannabinoids in anxiety responses: CB1 and GABAB receptors in the balance of GABAergic and glutamatergic neurotransmission.Neuropsychopharmacology. 2012;37(12): 2624–34.PubMed Abstract |Publisher Full Text |Free Full Text
- 36. Dubreucq S, Durand A, Matias I, et al.: Ventral tegmental area cannabinoid type-1 receptors control voluntary exercise performance.Biol Psychiatry. 2013;73(9): 895–903.PubMed Abstract |Publisher Full Text
- 37. Fuss J, Steinle J, Bindila L, et al.: A runner's high depends on cannabinoid receptors in mice.Proc Natl Acad Sci U S A. 2015;112(42): 13105–8.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 38. Martín-García E, Bourgoin L, Cathala A, et al.: Differential Control of Cocaine Self-Administration by GABAergic and Glutamatergic CB1 Cannabinoid Receptors.Neuropsychopharmacology. 2015.PubMed Abstract |Publisher Full Text
- 39. Talani G, Lovinger DM: Interactions between ethanol and the endocannabinoid system at GABAergic synapses on basolateral amygdala principal neurons.Alcohol. 2015;49(8): 781–94.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 40. Albayram Ö, Passlick S, Bilkei-Gorzo A, et al.: Physiological impact of CB1 receptor expression by hippocampal GABAergic interneurons.Pflugers Arch. 2016;468(4): 727–37.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 41. Puighermanal E, Marsicano G, Busquets-Garcia A, et al.: Cannabinoid modulation of hippocampal long-term memory is mediated by mTOR signaling.Nat Neurosci. 2009;12(9): 1152–8.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 42. Chiarlone A, Bellocchio L, Blázquez C, et al.: A restricted population of CB1 cannabinoid receptors with neuroprotective activity.Proc Natl Acad Sci U S A. 2014;111(22): 8257–62.PubMed Abstract |Publisher Full Text
- 43. Metna-Laurent M, Soria-Gómez E, Verrier D, et al.: Bimodal control of fear-coping strategies by CB1 cannabinoid receptors.J Neurosci. 2012;32(21): 7109–18.PubMed Abstract |Publisher Full Text
- 44. Terzian AL, Micale V, Wotjak CT: Cannabinoid receptor type 1 receptors on GABAergic vs. glutamatergic neurons differentially gate sex-dependent social interest in mice.Eur J Neurosci. 2014;40(1): 2293–8.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 45. Dubreucq S, Matias I, Cardinal P, et al.: Genetic dissection of the role of cannabinoid type-1 receptors in the emotional consequences of repeated social stress in mice.Neuropsychopharmacology. 2012;37(8): 1885–900.PubMed Abstract |Publisher Full Text
- 46. Han J, Kesner P, Metna-Laurent M, et al.: Acute cannabinoids impair working memory through astroglial CB1 receptor modulation of hippocampal LTD.Cell. 2012;148(5): 1039–50.PubMed Abstract |Publisher Full Text
- 47. Molina-Holgado E, Vela JM, Arévalo-Martín A, et al.: Cannabinoids promote oligodendrocyte progenitor survival: involvement of cannabinoid receptors and phosphatidylinositol-3 kinase/Akt signaling.J Neurosci. 2002;22(22): 9742–53.PubMed Abstract
- 48. Stella N: Endocannabinoid signaling in microglial cells.Neuropharmacology. 2009;56(Suppl 1): 244–53.PubMed Abstract |Publisher Full Text |Free Full Text
- 49. Mecha M, Feliú A, Carrillo-Salinas FJ, et al.: Endocannabinoids drive the acquisition of an alternative phenotype in microglia.Brain Behav Immun. 2015;49: 233–45.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 50. Oliveira da Cruz JF, Robin LM, Drago F, et al.: Astroglial type-1 cannabinoid receptor (CB1): A new player in the tripartite synapse.Neuroscience. 2016;323: 35–42.PubMed Abstract |Publisher Full Text
- 51. Metna-Laurent M, Marsicano G: Rising stars: modulation of brain functions by astroglial type-1 cannabinoid receptors.Glia. 2015;63(3): 353–64.PubMed Abstract |Publisher Full Text
- 52. Navarrete M, Díez A, Araque A: Astrocytes in endocannabinoid signalling.Philos Trans R Soc Lond B Biol Sci. 2014;369(1654): 20130599.PubMed Abstract |Publisher Full Text |Free Full Text
- 53. Pérez-Alvarez A, Araque A: Astrocyte-neuron interaction at tripartite synapses.Curr Drug Targets. 2013;14(11): 1220–4.PubMed Abstract |Publisher Full Text
- 54. Martín R, Bajo-Grañeras R, Moratalla R, et al.: Circuit-specific signaling in astrocyte-neuron networks in basal ganglia pathways.Science. 2015;349(6249): 730–4.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 55. Oliveira JF, Sardinha VM, Guerra-Gomes S, et al.: Do stars govern our actions? Astrocyte involvement in rodent behavior.Trends Neurosci. 2015;38(9): 535–49.PubMed Abstract |Publisher Full Text
- 56. Dudok B, Barna L, Ledri M, et al.: Cell-specific STORM super-resolution imaging reveals nanoscale organization of cannabinoid signaling.Nat Neurosci. 2015;18(1): 75–86.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 57. Thibault K, Carrel D, Bonnard D, et al.: Activation-dependent subcellular distribution patterns of CB1 cannabinoid receptors in the rat forebrain.Cereb Cortex. 2013;23(11): 2581–91.PubMed Abstract |Publisher Full Text
- 58. Bénard G, Massa F, Puente N, et al.: Mitochondrial CB1 receptors regulate neuronal energy metabolism.Nat Neurosci. 2012;15(4): 558–64.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 59. Howlett AC: The cannabinoid receptors.Prostaglandins Other Lipid Mediat. 2002;68–69: 619–31.PubMed Abstract
- 60. Bartova A, Birmingham MK: Effect of delta9-tetrahydrocannabinol on mitochondrial NADH-oxidase activity.J Biol Chem. 1976;251(16): 5002–6.PubMed Abstract
- 61. Aquila S, Guido C, Santoro A, et al.: Rimonabant (SR141716) induces metabolism and acquisition of fertilizing ability in human sperm.Br J Pharmacol. 2010;159(4): 831–41.PubMed Abstract |Publisher Full Text |Free Full Text
- 62. Arrabal S, Lucena MA, Canduela MJ, et al.: Pharmacological Blockade of Cannabinoid CB1 Receptors in Diet-Induced Obesity Regulates Mitochondrial Dihydrolipoamide Dehydrogenase in Muscle.PLoS One. 2015;10(12): e0145244.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 63. Vallée M, Vitiello S, Bellocchio L, et al.: Pregnenolone can protect the brain from cannabis intoxication.Science. 2014;343(6166): 94–8.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 64. Hebert-Chatelain E, Reguero L, Puente N, et al.: Cannabinoid control of brain bioenergetics: Exploring the subcellular localization of the CB1 receptor.Mol Metab. 2014;3(4): 495–504.PubMed Abstract |Publisher Full Text |Free Full Text
- 65. Koch M, Varela L, Kim JG, et al.: Hypothalamic POMC neurons promote cannabinoid-induced feeding.Nature. 2015;519(7541): 45–50.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 66. Hollis F, van der Kooij MA, Zanoletti O, et al.: Mitochondrial function in the brain links anxiety with social subordination.Proc Natl Acad Sci U S A. 2015;112(50): 15486–91.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 67. Cota D, Marsicano G, Tschöp M, et al.: The endogenous cannabinoid system affects energy balance via central orexigenic drive and peripheral lipogenesis.J Clin Invest. 2003;112(3): 423–31.PubMed Abstract |Publisher Full Text |Free Full Text
- 68. Izzo AA, Sharkey KA: Cannabinoids and the gut: new developments and emerging concepts.Pharmacol Ther. 2010;126(1): 21–38.PubMed Abstract |Publisher Full Text
- 69. Jyotaki M, Shigemura N, Ninomiya Y: Modulation of sweet taste sensitivity by orexigenic and anorexigenic factors.Endocr J. 2010;57(6): 467–75.PubMed Abstract |Publisher Full Text
- 70. Nadolska K, Goś R: [The role of endocannabinoid system in physiological and pathological processes in the eye].Klin Oczna. 2008;110(10–12): 392–6.PubMed Abstract
- 71. Mach F, Steffens S: The role of the endocannabinoid system in atherosclerosis.J Neuroendocrinol. 2008;20(Suppl 1): 53–7.PubMed Abstract |Publisher Full Text
- 72. Osei-Hyiaman D, DePetrillo M, Pacher P, et al.: Endocannabinoid activation at hepatic CB1 receptors stimulates fatty acid synthesis and contributes to diet-induced obesity.J Clin Invest. 2005;115(5): 1298–305.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 73. Cota D: CB1 receptors: emerging evidence for central and peripheral mechanisms that regulate energy balance, metabolism, and cardiovascular health.Diabetes Metab Res Rev. 2007;23(7): 507–17.PubMed Abstract |Publisher Full Text
- 74. Jean-Gilles L, Braitch M, Latif ML, et al.: Effects of pro-inflammatory cytokines on cannabinoid CB1 and CB2 receptors in immune cells.Acta Physiol (Oxf). 2015;214(1): 63–74.PubMed Abstract |Publisher Full Text |Free Full Text
- 75. Idris AI, van 't Hof RJ, Greig IR, et al.: Regulation of bone mass, bone loss and osteoclast activity by cannabinoid receptors.Nat Med. 2005;11(7): 774–9.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 76. Bíró T, Tóth BI, Haskó G, et al.: The endocannabinoid system of the skin in health and disease: novel perspectives and therapeutic opportunities.Trends Pharmacol Sci. 2009;30(8): 411–20.PubMed Abstract |Publisher Full Text |Free Full Text
- 77. Cavuoto P, McAinch AJ, Hatzinikolas G, et al.: The expression of receptors for endocannabinoids in human and rodent skeletal muscle.Biochem Biophys Res Commun. 2007;364(1): 105–10.PubMed Abstract |Publisher Full Text
- 78. Maccarrone M, Bab I, Bíró T, et al.: Endocannabinoid signaling at the periphery: 50 years after THC.Trends Pharmacol Sci. 2015;36(5): 277–96.PubMed Abstract |Publisher Full Text |Free Full Text
- 79. Bellocchio L, Soria-Gómez E, Quarta C, et al.: Activation of the sympathetic nervous system mediates hypophagic and anxiety-like effects of CB1 receptor blockade.Proc Natl Acad Sci U S A. 2013;110(12): 4786–91.PubMed Abstract |Publisher Full Text |Free Full Text
- 80. Hofer SC, Ralvenius WT, Gachet MS, et al.: Localization and production of peptide endocannabinoids in the rodent CNS and adrenal medulla.Neuropharmacology. 2015;98: 78–89.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 81. Cani PD, Plovier H, Van Hul M, et al.: Endocannabinoids - at the crossroads between the gut microbiota and host metabolism.Nat Rev Endocrinol. 2016;12(3): 133–43.PubMed Abstract |Publisher Full Text
- 82. Geurts L, Everard A, Van Hul M, et al.: Adipose tissue NAPE-PLD controls fat mass development by altering the browning process and gut microbiota.Nat Commun. 2015;6: 6495.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 83. Vemuri VK, Makriyannis A: Medicinal chemistry of cannabinoids.Clin Pharmacol Ther. 2015;97(6): 553–8.PubMed Abstract |Publisher Full Text
- 84. Pertwee RG: GPR55: a new member of the cannabinoid receptor clan?Br J Pharmacol. 2007;152(7): 984–6.PubMed Abstract |Publisher Full Text |Free Full Text
- 85. Lauckner JE, Jensen JB, Chen HY, et al.: GPR55 is a cannabinoid receptor that increases intracellular calcium and inhibits M current.Proc Natl Acad Sci U S A. 2008;105(7): 2699–704.PubMed Abstract |Publisher Full Text |Free Full Text
- 86. Xiong W, Cui T, Cheng K, et al.: Cannabinoids suppress inflammatory and neuropathic pain by targeting α3 glycine receptors.J Exp Med. 2012;209(6): 1121–34.PubMed Abstract |Publisher Full Text |Free Full Text
- 87. Di Marzo V, De Petrocellis L: Why do cannabinoid receptors have more than one endogenous ligand?Philos Trans R Soc Lond B Biol Sci. 2012;367(1607): 3216–28.PubMed Abstract |Publisher Full Text |Free Full Text
- 88. Christensen R, Kristensen PK, Bartels EM, et al.: Efficacy and safety of the weight-loss drug rimonabant: a meta-analysis of randomised trials.Lancet. 2007;370(9600): 1706–13.PubMed Abstract |Publisher Full Text
- 89. Volkow ND, Baler RD, Compton WM, et al.: Adverse health effects of marijuana use.N Engl J Med. 2014;370(23): 2219–27.PubMed Abstract |Publisher Full Text |Free Full Text
- 90. Janero DR: Cannabinoid-1 receptor (CB1R) blockers as medicines: beyond obesity and cardiometabolic disorders to substance abuse/drug addiction with CB1R neutral antagonists.Expert Opin Emerg Drugs. 2012;17(1): 17–29.PubMed Abstract |Publisher Full Text
- 91. Whiting PF, Wolff RF, Deshpande S, et al.: Cannabinoids for Medical Use: A Systematic Review and Meta-analysis.JAMA. 2015;313(24): 2456–73.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 92. Gentry PR, Sexton PM, Christopoulos A: Novel Allosteric Modulators of G Protein-coupled Receptors.J Biol Chem. 2015;290(32): 19478–88.PubMed Abstract |Publisher Full Text |Free Full Text
- 93. van der Westhuizen ET, Valant C, Sexton PM, et al.: Endogenous allosteric modulators of G protein-coupled receptors.J Pharmacol Exp Ther. 2015;353(2): 246–60.PubMed Abstract |Publisher Full Text
- 94. Price MR, Baillie GL, Thomas A, et al.: Allosteric modulation of the cannabinoid CB1 receptor.Mol Pharmacol. 2005;68(5): 1484–95.PubMed Abstract |Publisher Full Text
- 95. Horswill JG, Bali U, Shaaban S, et al.: PSNCBAM-1, a novel allosteric antagonist at cannabinoid CB1 receptors with hypophagic effects in rats.Br J Pharmacol. 2007;152(5): 805–14.PubMed Abstract |Publisher Full Text |Free Full Text
- 96. Navarro HA, Howard JL, Pollard GT, et al.: Positive allosteric modulation of the human cannabinoid (CB) receptor by RTI-371, a selective inhibitor of the dopamine transporter.Br J Pharmacol. 2009;156(7): 1178–84.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 97. Abood ME: Allosteric Modulators: A Side Door.J Med Chem. 2016;59(1): 42–3.PubMed Abstract |Publisher Full Text
- 98. Morales P, Goya P, Jagerovic N, et al.: Allosteric Modulators of the CB1 Cannabinoid Receptor: A Structural Update Review.Cannabis and Cannabinoid Research. 2016;1(1): 22–30.Publisher Full Text
- 99. Ignatowska-Jankowska BM, Baillie GL, Kinsey S, et al.: A Cannabinoid CB1 Receptor-Positive Allosteric Modulator Reduces Neuropathic Pain in the Mouse with No Psychoactive Effects.Neuropsychopharmacology. 2015;40(13): 2948–59.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 100. Pamplona FA, Ferreira J, Menezes de Lima O Jr, et al.: Anti-inflammatory lipoxin A4 is an endogenous allosteric enhancer of CB1 cannabinoid receptor.Proc Natl Acad Sci U S A. 2012;109(51): 21134–9.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 101. Laprairie RB, Bagher AM, Kelly ME, et al.: Cannabidiol is a negative allosteric modulator of the cannabinoid CB1 receptor.Br J Pharmacol. 2015;172(20): 4790–805.PubMed Abstract |Publisher Full Text |Free Full Text
- 102. Hill AJ, Williams CM, Whalley BJ, et al.: Phytocannabinoids as novel therapeutic agents in CNS disorders.Pharmacol Ther. 2012;133(1): 79–97.PubMed Abstract |Publisher Full Text
- 103. Chiodi V, Ferrante A, Ferraro L, et al.: Striatal adenosine-cannabinoid receptor interactions in rats over-expressing adenosine A2A receptors.J Neurochem. 2016;136(5): 907–17.PubMed Abstract |Publisher Full Text
- 104. Viñals X, Moreno E, Lanfumey L, et al.: Cognitive Impairment Induced by Delta9-tetrahydrocannabinol Occurs through Heteromers between Cannabinoid CB1 and Serotonin 5-HT2A Receptors.PLoS Biol. 2015;13(7): e1002194.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 105. Callén L, Moreno E, Barroso-Chinea P, et al.: Cannabinoid receptors CB1 and CB2 form functional heteromers in brain.J Biol Chem. 2012;287(25): 20851–65.PubMed Abstract |Publisher Full Text |Free Full Text
- 106. Maccarrone M, Bernardi G, Agrò AF, et al.: Cannabinoid receptor signalling in neurodegenerative diseases: a potential role for membrane fluidity disturbance.Br J Pharmacol. 2011;163(7): 1379–90.PubMed Abstract |Publisher Full Text |Free Full Text
- 107. Lee SH, Ledri M, Tóth B, et al.: Multiple Forms of Endocannabinoid and Endovanilloid Signaling Regulate the Tonic Control of GABA Release.J Neurosci. 2015;35(27): 10039–57.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 108. Zhang HY, Gao M, Shen H, et al.: Expression of functional cannabinoid CB2 receptor in VTA dopamine neurons in rats.Addict Biol. 2016.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 109. Li Y, Kim J: CB2 Cannabinoid Receptor Knockout in Mice Impairs Contextual Long-Term Memory and Enhances Spatial Working Memory.Neural Plast. 2016;2016: 9817089.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 110. Li Y, Kim J: Deletion of CB2 cannabinoid receptors reduces synaptic transmission and long-term potentiation in the mouse hippocampus.Hippocampus. 2016;26(3): 275–81.PubMed Abstract |Publisher Full Text |Faculty Opinions Recommendation
- 111. Pistis M, Melis M: From surface to nuclear receptors: the endocannabinoid family extends its assets.Curr Med Chem. 2010;17(14): 1450–67.PubMed Abstract |Publisher Full Text
- 112. Fezza F, Bari M, Florio R, et al.: Endocannabinoids, related compounds and their metabolic routes.Molecules. 2014;19(11): 17078–106.PubMed Abstract |Publisher Full Text
- 113. Viader A, Blankman JL, Zhong P, et al.: Metabolic Interplay between Astrocytes and Neurons Regulates Endocannabinoid Action.Cell Rep. 2015;12(5): 798–808.PubMed Abstract |Publisher Full Text |Free Full Text |Faculty Opinions Recommendation
- 114. Viader A, Ogasawara D, Joslyn CM, et al.: A chemical proteomic atlas of brain serine hydrolases identifies cell type-specific pathways regulating neuroinflammation.eLife. 2016;5: e12345.PubMed Abstract |Publisher Full Text |Free Full Text
- 115. Steindel F, Lerner R, Häring M, et al.: Neuron-type specific cannabinoid-mediated G protein signalling in mouse hippocampus.J Neurochem. 2013;124(6): 795–807.PubMed Abstract |Publisher Full Text
- 116. Bellocchio: Patent: Compositions targeting cb1 receptor for controlling food intake. Application Number: EP09306163.8. European Patent Office. 2010.
- 117. Di Marzo V, Stella N, Zimmer A: Endocannabinoid signalling and the deteriorating brain.Nat Rev Neurosci. 2015;16(1): 30–42.PubMed Abstract |Publisher Full Text |Free Full Text


Comments on this articleComments (0)